Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Биосинтез углеводов в животных тканях
Краткое содержание главы
Глюконеогенез - это образование «нового» сахара из неуглеводных предшественников, среди которых наибольшее значение имеют пиру ват, лактат, промежуточные продукты цикла лимонной кислоты и многие аминокислоты. Подобно всем прочим биосинтетическим путям, ферментативный путь глюконеогенеза не идентичен соответствующему катаболическому пути, регулируется независимо от него и требует расхода химической энергии в форме АТР. Синтез глюкозы из пирувата происходит у позвоночных главным образом в печени и отчасти в почках. На этом биосинтетическом пути используются семь ферментов, участвующих в гликолизе; они функционируют обратимо и присутствуют в большом избытке. Однако на гликолитическом пути, т.е. на пути «вниз», имеются также три необратимые стадии, которые не могут использоваться в глюконеогенезе. В этих пунктах глюконеогенез идет в обход гликолитического пути, за счет других реакций, катализируемых другими ферментами. Первый обходный путь - это превращение пирувата в фосфоенолпируват через оксалоацетат; второй - это дефосфорилирование фруктозо-1,6-дифосфата, катализируемое фруктозодифосфатазой, и, наконец, третий обходный путь - это дефосфорилирование глюкозо-6-фосфата, катализируемое глюкозо-6-фосфатазой. На каждую молекулу D-глюкозы, образующуюся из пирувата, расходуются концевые фосфатные группы четырех молекул АТР и двух молекул GTP. Регулируется глюконеогенез через две главные стадии: 1) карбоксилирование пирувата, катализируемое пируваткарбоксилазой, которая активируется аллостерическим эффектором ацетил-СоА, и 2) дефосфорилирование фруктозо-1,6-дифосфата, катализируемое фруктозодифосфатазой, которая ингибируется АМР и активируется цитратом. По три атома углерода от каждого промежуточного продукта цикла лимонной кислоты и углеродные скелеты многих аминокислот способны превращаться в глюкозу. Из жирных кислот с четным числом атомов углерода и из ацетил-СоА реального образования глюкозы не происходит, тогда как три углеродных атома жирных кислот с нечетным числом атомов углерода, а также образуемый бактериями рубца пропионат могут превращаться в глюкозу: при этом в качестве промежуточного продукта образуется метилмалонил-СоА, превращающийся затем в сукцинил-СоА при участии кофермента В12. В периоды восстановления после напряженной мышечной работы глюконеогенез протекает очень активно, благодаря чему присутствующий в крови лактат превращается в гликоген и глюкозу.
Путь синтеза гликогена также отличается от пути, по которому идет его расщепление. Он включает превращение глюкозо-1-фосфата в уридиндифосфат-глюкозу, которая затем - при участии гликоген-синтазы - передает глюкозильные группы на нередуцирующий конец боковых цепей гликогена. Новые боковые цепи возникают в молекулах гликогена в результате действия гликозил-(46)-трансферазы [а(1,4→1,6)- трансгликозилазы]. Процессы синтеза и расщепления гликогена регулируются независимо и репипрокно. Соотношение скоростей этих двух процессов контролируется гормонами адреналином и глюкагоном. Известен ряд генетических дефектов, при которых синтез или расщепление гликогена нарушены.
Синтез лактозы в молочной железе происходит при участии лактальбумингалактозилтрансферазного комплекса. Лактальбумин в этом комплексе выполняет роль субъединицы, изменяющей специфичность фермента. Его образование регулируется гормонами, вызывающими лактацию.
ЛИТЕРАТУРА
Книги
Cmninggham Е.В. Biochemistry: Mechanisms of Metabolism, McGraw-Hill, New York, 1978.
В гл. 9 содержатся дополнительные сведения, касающиеся биосинтеза углеводов, главным образом энзимологические данные.
Dickens F., Randle P.J., Whelan W.J. (eds.). Carbohydrate Metabolism and Its Disorders, vols. I and II, Academic, New York. 1968. Сборник ценных обзоров.
Newsholme E. A., Start C. Regulation in Metabolism, Wiley, New York, 1973. Гл. 4 и 6 включают более подробные данные по регуляции глюконеогенеза и синтеза гликогена.
Статьи
Bent Н. A. Energy and Exercise, J. Chem. Ed., vol. 55, nos. 7-12, July-December 1978.
Howell R. R. The Glycogen Storage Diseases, pp. 160-181. In: J.B. Stanbury, J. B. Wyngaarden and D. S. Frederickson (eds.), The Metabolic Basis of Inherited Disease, 4th ed, McGraw-Hill, New York, 1978.
Katz J., Rognstad R., Futile Cycles in the Metabolism of Glucose, Curr. Top. Cell Regul., 10, 238-287 (1976).
Krebs H.A. Some Aspects of the Regulation of Fuel Supply in Omnivorous Animals, Adv. Enz. Regul., 10, 397-420 (1972).
Sharon N. Carbohydrates, Sсi. Am, 243, 90-116, November 1980.
Вопросы и задачи
1. Роль окислительного фосфорилирования в глюконеогенезе. Возможен ли реальный синтез глюкозы из пирувата в условиях, когда цикл лимонной кислоты и окислительное фосфорилирование полностью ингибированы?
2. Путь атомов углерода в глюконеогенезе. Печеночный экстракт, способный катализировать все обычные метаболические реакции, инкубируют непродолжительное время в отдельных опытах со следующими двумя предшественниками, меченными 14С:
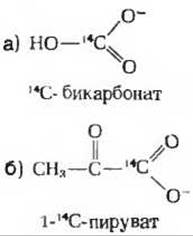
Проследите путь каждого из этих двух предшественников в глюконеогенезе. В каком положении обнаружится метка во всех промежуточных продуктах и в конечном продукте, т. е. в глюкозе?
3. Путь СО2 в глюконеогенезе. В первой обходной реакции глюконеогенеза превращении пирувата в фосфоенолпируват сначала под действием пируваткарбоксилазы пируват карбоксилируется с образованием оксалоацетата а затем оксало- ацетат декарбоксилируется до фосфоенолпирувата в реакции, катализируемой фосфоенолпируват-карбоксикиназой. Поскольку за присоединением СО2 непосредственно следует ее отщепление, можно было бы думать, что 14С из 14СО2 не будет включаться в фосфоенолпируват и глюкозу, равно как и в любой другой из промежуточных продуктов глюконеогенеза. Выяснилось, однако, что если срезы крысиной печени синтезируют глюкозу в присутствии 14СО2, то 14С хотя и не сразу, но все же обнаруживается в фосфоенолпирувате и в конце концов - в 3-м и 4-м углеродных атомах глюкозы. Каким образом метка попадает в фосфоенолпируват и в 3-й и 4-й углеродные атомы глюкозы? (Подсказка: если глюконеогенез происходит в присутствии 14СО2. то некоторые четырехуглеродные промежу очные продукты цикла лимонной кислоты тоже оказываются мечеными.)
4. Регуляция фруктозодифюсфатазы и фосфофруктокиназы. Как влияет повышение концентраций АТР и АМР на каталитическую активность фруктозодифосфагазы и фосфофруктокиназы? Как сказываются эти эффекты на относительной величине потока метаболитов в глюконеогенезе и гликолизе?
5. Глюкогенные субстраты. Чтобы определить, может ли то или иное соединение служить предшественником глюкозы, поступают обычно следующим образом: сначала оставляют животное голодать, пока у него не истощится запас гликогена в печени, а потом дают ему исследуемое соединение. Те соединения, под влиянием которых количество гликогена в печени увеличивается, принято называть глюкогенными, потому что они сначала превращаются в глюкозо-6-фосфат. Ниже приведены формулы некоторых соединений. Покажите на основе известных ферментативных реакций, какие из них являются глюкогенными.
6. Уровень лактата в крови при большой физической нагрузке. На рисунке показана концентрация лактата в крови до бега на 400 м. во время бега и после него.
Задача 6
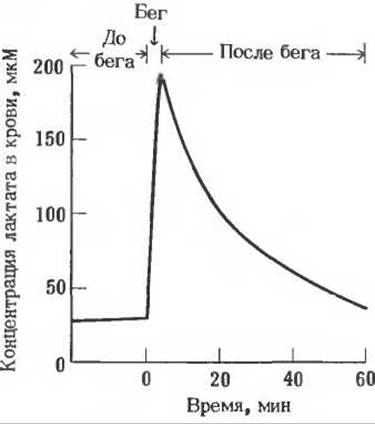
а) Чем вызывается быстрое повышение концентрации лактата?
б) Что является причиной снижения уровня лактата после бега? Почему снижение происходит медленнее, чем подъем?
в) Почему в состоянии покоя концентрация лактата в крови не равна нулю?
7. Избыточное потребление кислорода во время глюконеогенеза. Поглощаемый печенью лактат превращается в глюкозу. Этот процесс требует затраты АТР; 6 молекул АТР расходуются на образование 1 молекулы глюкозы. Об интенсивности этого процесса в срезах крысиной печени можно судить, вводя в среду 14С-лактат и измеряя количество образовавшейся 14С-глюкозы. Поскольку стехиометрические соотношенич между потреблением О2 и синтезом АТР известны (гл. 17), мы можем предсказать, какое дополнительное количество кислорода (сверх обычного уровня) должно быть потреблено при введении данного количества лактата. Фактические измерения показывают, однако, что этот избыток О2, необходимый для образования глюкозы из лактата, всегда выше предсказанного на основе известных стехиометрических соотношений. Предложите возможное объяснение этого факта.
8. В какой точке регулируется синтез гликогена? Объясните, каким образом два приведенных ниже наблюдения могут помочь определить регулируемую стадию синтеза гликогена в скелетных мышцах?
а) Измеренная активность гликоген-синтазы в покоящейся мышце (выраженная в микромолях UDP-глюкозы, использованной на грамм в минуту) ниже, чем активность фосфоглюкомутазы или UDP-глюкозопирофосфорилазы (выраженная в микромолях субстрата, переработанного на грамм в минуту).
б) Стимуляция синтеза гликогена вызывает незначительное снижение концентраций глюкозо-6-фосфата и глюкозо-1-фосфата, резкое падение концентрации UDP-глюкозы и существенное увеличение концентрации UDP.
9. Во что обходится организму хранение глюкозы в форме гликогена? Напишите ряд последовательных реакций и суммарную реакцию, которые позволят определить число молекул АТР, расходуемых на превращение цитоплазматического глюкозо-6-фосфата в гликоген и гликогена обратно в глюкозо-6-фосфат. Какую часть это число составит от максимального числа молекул АТР, образующегося при полном расщеплении глюкозо-6-фосфата?
10. Выявление дефектных ферментов углеводного обмена. Проводилось патолого-анатомическое исследование кусочка ткани печени. У больного подозревали генетический дефект одного из ферментов углеводного обмена. Было показано, что гомогенат ткани 1) расщеплял гликоген до глюкозо-6-фосфата, 2) не обладал способностью синтезировать гликоген из какого бы то ни было сахара, так же как и способностью использовать галактозу в качестве источника энергии, и 3) синтезировал глюкозо-6-фосфат из лактата. Ниже названы три фермента; какой из них был дефектным в этом случае:
а) гликоген-фосфорилаза,
б) фруктозодифосфатаза,
в) UDP-глюкозопирофосфорилаза? Аргументируйте свой выбор.
11. Кетоз у овец. Около 80% всей глюкозы, синтезируемой в организме овцы, используется в вымени. Расходуется глюкоза на образование молока, главным образом на синтез лактозы, а также глицеролфосфата, необходимого для синтеза триацилглицеролов молока. Зимой при низком качестве корма образуется меньше молока и у овец иногда развивается кетоз, т.е. повышается концентрация кетоновых тел в плазме крови. Из-за чего происходят эти изменения? Обычно животным в таких случаях дают большие дозы пропионата. На чем основано его действие?
12. Адаптация к галактоземии. Галактоземия - это патологическое состояние, в основе которого лежит неспособность к утилизации галактозы, образующейся из лактозы, содержащейся в пище. Одна из форм этою заболевания обусловлена отсутствием фермента, называемого галактозо-1-фосфат - уридилтрансферазой. Если ребенок с таким дефектом не умирает в раннем возрасте, то позже у него может в какой-то мере развиться способность метаболизировать галактозу. Эта способность появляется за счет усиленного образования фермента UDP-галактозопирофосфорилазы, катализирующего следующую реакцию:
![]()
Каким образом благодаря присутствию этого фермента у таких больных развивается способность метаболизировать галактозу?