Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Гликолиз - центральный путь катаболизма глюкозы
Пути, ведущие от гликогена и других углеводов, к центральному гликолитическому пути
Не только D-глюкоза, но и многие другие углеводы вовлекаются после ряда превращений в гликолиз, высвобождая заключенную в них энергию. Среди этих углеводов главную роль играют запасные полисахариды - гликоген и крахмал, дисахариды - мальтоза, лактоза и сахароза и моносахариды - фруктоза, манноза и галактоза. Пути, по которым эти различные углеводы вступают на путь гликолиза, показаны на рис. 15-8.
D-глюкозные единицы боковых цепей гликогена и крахмала вовлекаются в гликолиз в результате последовательного действия двух ферментов - гликоген-фосфорилазы (или фосфорилазы крахмала у растений) и фосфоглюкомутазы. Гликоген-фосфорилаза, широко распространенная в животных клетках, катализирует изображенную ниже общую реакцию, в которой (Глюкоза), означает боковую цепь гликогена (или крахмала), состоящую из n D-глюкозных единиц, соединенных а(1→4)-связями, а (Глюкоза)n-1 - ту же боковую цепь, но только укороченную на одно звено в результате отщепления от ее конца одного остатка глюкозы (структуру гликогена и крахмала см. на рис. 11-15):
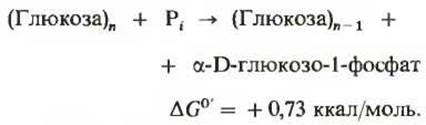
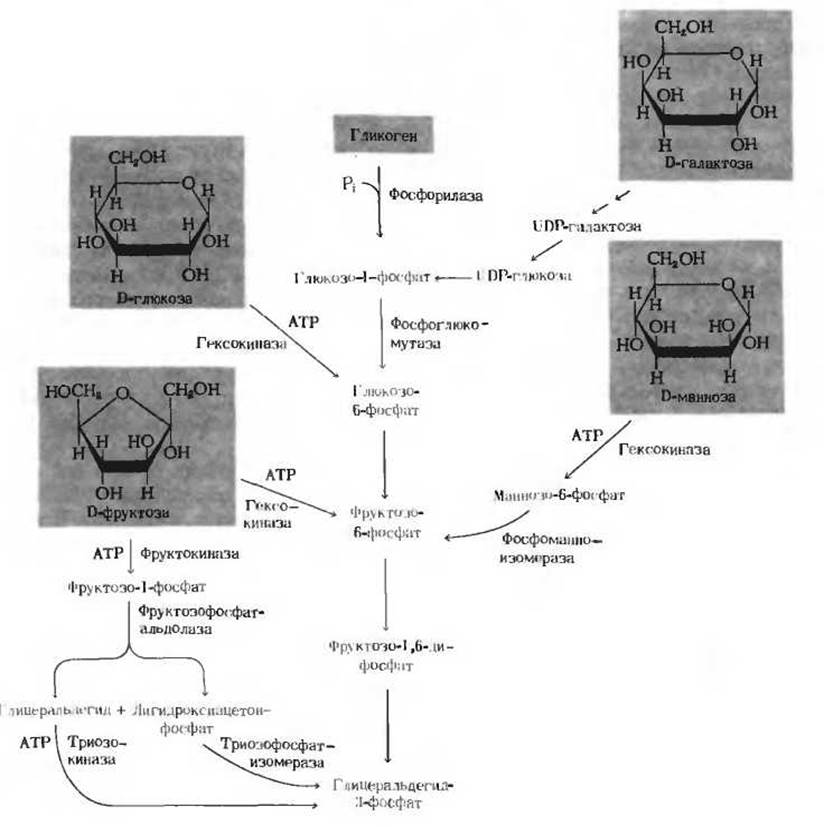
Рис. 15-8. Вовлечение гликогена и различных гексоз в первую стадию гликолиза.
В условиях клетки, при относительно высокой концентрации фосфата, гликогенфосфорилазная реакция идет только в сторону распада гликогена и образования глюкозо-1-фосфата. В этой реакции концевая а (1→4)-гликозидная связь на нередуцирующем конце боковой цепи гликогена претерпевает фосфорелиз: под действием фосфата от цепи отщепляется концевой остаток глюкозы с образованием a-D-глюкозо-1-фосфата. Боковая цепь гликогена становится в результате этой реакции короче на одну глюкозную единицу (рис. 15-9). Гликоген-фосфорилаза атакует нередуцирующие концы боковых цепей гликогена многократно, до тех пор пока она не дойдет до точки, отстоящей на четыре глюкозные единицы от а (1→6)-связи (рис. 11-15). Здесь ее действие прекращается.
Для того чтобы расщепление гликогена под действием гликогенфосфорилазы могло продолжаться, на полисахарид должен предварительно подействовать другой фермент, а (1→6)-глюкозидаза. Этот фермент катализирует две реакции. В первой из них он отщепляет от цепи три глюкозных остатка из упомянутых четырех и переносит их на конец какой-нибудь другой внешней боковой цепи. Во второй реакции, катализируемой а (1→6)-глюкозидазой, отщепляется четвертый глюкозный остаток, присоединенный в точке ветвления a(1→6)-связью. Гидролиз а(1→6)-связи в точке ветвления приводит к образованию одной молекулы D-глюкозы и открывает для действия гликоген-фосфорилазы новый участок цепи, состоящий из остатков глюкозы, соединенных а (1→4)-связями.
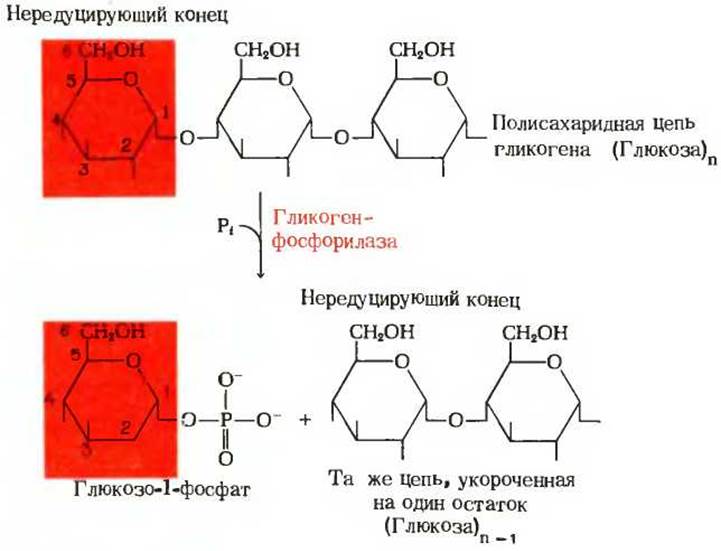
Рис. 15-9. Удаление концевого остатка глюкозы на нередуцирующем конце одной из цепей гликогена под действием гликогенфосфорилазы. Этот процесс многократно повторяется, и остатки глюкозы отщепляются один за другим до тех пор, пока концевым не окажется остаток, четвертый по счету от точки ветвления (см. текст). Обратите внимание на условное обозначение гидроксильных групп остатков глюкозы; водородные атомы, присоединенные к пиранозным кольцам, не показаны.
Глюкозо-1-фосфат - конечный продукт реакции, катализируемой гликоген-фосфорилазой (фосфорилазой крахмала), превращается в глюкозо-6-фосфат под действием фермента фосфоглюкомутазы. Этот фермент (он был выделен в чистом виде из многих источников) катализирует обратимую реакцию:
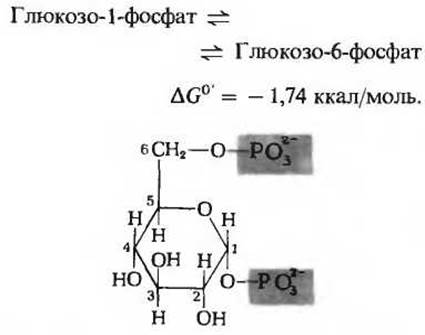
Рис. 15-10. a-D-глюкозо-1,6-дифосфат - один из кофакторов, необходимых для действия фосфоглюкомутазы.
Для действия фосфоглюкомутазы необходим в качестве кофактора глюкозо-1,6-дифосфат (рис. 15-10). Роль этого кофактора станет ясной, если учесть промежуточные этапы действия фермента. Из приведенной ниже последовательности реакций видно, что фермент пребывает попеременно в одной из двух форм - фосфорилированной и нефосфорилированной, или дефосфорилированной:
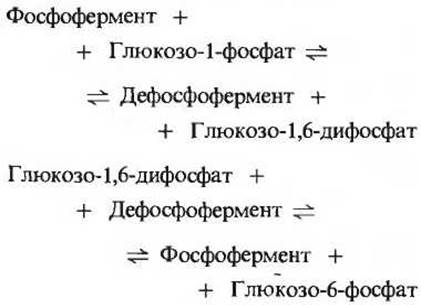
Суммарное уравнение:
![]()
Фосфоглюкомутаза примечательна и еще в одном отношении: этот фермент является представителем обширного класса ферментов, у которых в активном центре присутствует остаток серина, необходимый для каталитической активности. Именно этот остаток серина участвует во взаимодействии с глюкозо-1,6-дифосфатом - его гидроксильная группа этерифицируется фосфорной кислотой. Ферменты серинового класса (рис. 9-12), к которым принадлежит и фосфоглюкомутаза, необратимо ингибируются некоторыми органическими фосфатами, такими, как, например, диизопропилфторфосфат. При этом ингибиторы взаимодействуют с гидроксильной группой упомянутого остатка серина, в результате чего образуется фосфорилированное производное фермента, лишенное каталитической активности (рис. 9-10).