Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
Бактериальные экстракты содержат ДНК-полимеразу
Теперь, когда мы получили общее представление о репликации ДНК, рассмотрим вопрос об участии ферментов в этом процессе. Впервые механизмы действия ферментов при репликации стали доступны прямому биохимическому исследованию благодаря важным работам Артура Корнберга и его коллег, начатым в 1956 г. Они инкубировали экстракты из клеток Е. coli со смесью dATP, dTTP, dGTP и dCTP, а-фосфатная группа которых была помечена изотопом 32Р (рис. 28-6). Было обнаружено, что при этом синтезируется очень небольшое количество новой ДНК, содержащей в своих фосфатных группах изотоп 32Р. Фермент, катализирующий эту реакцию и названный ДНК-полимеразой I, был в конце концов очищен и его свойства были подробно изучены. Оказалось, что он катализирует последовательное присоединение дезоксирибонуклеотидных остатков к концу цепи ДНК с одновременным высвобождением неорганического пирофосфата, содержащего ß- и у-фосфатные группы каждого встраивающегося дезоксирибонуклеозид-5'-трифосфата. Уравнение реакции в простейшей форме имеет вид
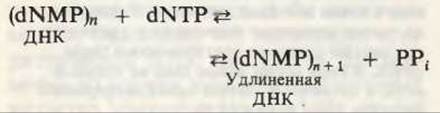
где dNMP и dNTP означают соответственно дезоксирибонуклеозид-5'-монофосфат и дезоксирибонуклеозид-5'-трифосфат. Если хотя бы один из четырех предшественников отсутствует, то новая ДНК не образуется, т.е. синтез новой ДНК идет только в присутствии всех четырех предшественников. 5'-трифосфаты всех четырех дезоксирибонуклеозидов не могут быть заменены соответствующими 5'-дифосфатами или 5'-монофосфатами; фермент не работает также с рибонуклеозид-5'-трифосфатами. Для работы ДНК-полимеразе необходимы ионы Mg2+, а в ее активном центре содержится прочно связанный с ферментом ион Zn2+.
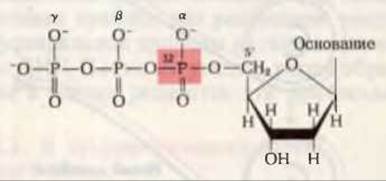
Рис. 28-6. Дезоксирибонуклеозид-5'-трифосфат, меченный [32Р] в а-положении.
ДНК-полимераза катализирует ковалентное связывание новых дезоксирибонуклеотидов, которое происходит благодаря присоединению их а-фосфатных групп к свободному 3'-гидроксильному концу предсуществующей цепи ДНК; следовательно, синтез цепи ДНК происходит в направлении 5'→3' (рис. 28-7). Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в остове ДНК, обеспечивается расщеплением пирофосфатной связи между а- и ß-фосфатными группами предшественников - дезоксирибонуклеозид-5'-трифосфатов. Образующийся при этом пирофосфат разрушается затем до фосфата, который может сдвигать реакцию в сторону ее завершения. В процессе работы было сделано очень важное наблюдение: было отмечено, что ДНК-полимеразная реакция протекает только в том случае, если в системе уже находится некоторое количество предсуществующей двухцепочечной ДНК.