Биохимия человека Том 1 - Марри Р. 1993
Метаболизм белков и аминокислот
Порфирины и желчные пигменты
Катаболизм гема: образование желчных пигментов
При физиологических условиях в организме взрослого человека разрушается 1—2∙108 эритроцитов в час. Таким образом, в течение суток у человека массой 70 кг обновляется приблизительно 6 г гемоглобина. При разрушении гемоглобина его белковая часть (глобин) может быть использована как таковая или после гидролиза в форме составляющих ее аминокислот; железо гема включается в общий пул железа и также снова используется. Вместе с тем свободная от железа порфириновая часть гема обязательно деградирует; это в основном происходит в ретикулоэндотелиальных клетках печени, селезенки и костного мозга.
Катаболизм гема, освобожденного из любых гемовых белков, осуществляется в микросомальной фракции ретикулоэндотелиальных клеток сложной ферментной системой — гем-оксигеназой. К моменту поступления гема из гемовых белков в гем-оксигеназную систему железо обычно окисляется в ферри-форму (гем превращается в гемин); гемин может легко связываться с альбумином с образованием метгемальбумина. Гем-оксигеназная система индуцируется субстратом. Она локализована около микросомальной системы транспорта электронов. Как показано на рис. 33.12, гемин восстанавливается в ферро-форму с помощью NADPH; далее при участии NADPH кислород присоединяется к а- метенильному мостику между пиррольными кольцами I и II. Ферро-форма железа снова окисляется в ферри-форму. При последующем присоединении кислорода происходит освобождение ферри-иона, выделение молекулы окиси углерода и образование в результате раскрытия тетрапиррольного кольца эквимолярного количества биливердина IХ-а. В этой реакции сам гем участвует в роли катализатора.

Рис. 33.12. Схема микросомальной гем-оксигеназной системы. (Воспроизведено с изменениями из обзора Schmid R., McDonough А. Е. The Porphyrins. Dolphin D. (ed.). Academic Press, 1978.)
У птиц и земноводных зеленый пигмент биливердин IХ-а экскретируется из организма; у млекопитающих растворимый фермент биливердинредуктаза катализирует восстановление метенильного мостика между пирролами III и IV в метиленовую группу, в результате образуется желтый пигмент билирубин ІХ-а (рис. 33.12).
По расчету из 1 г гемоглобина образуется 35 мг билирубина. Суточное образование билирубина у взрослого человека составляет приблизительно 250—350 мг.
Химическое превращение гема в билирубин ретикулоэндотелиальными клетками можно наблюдать in vivo: в гематоме обусловленный гемом пурпурный цвет медленно переходит в желтый цвет билирубина.
Дальнейшей метаболизм билирубина в основном происходит в печени. Он складывается из трех процессов: 1) поглощение билирубина паренхимальными клетками печени; 2) конъюгация билирубина в гладком эндоплазматическом ретикулуме и 3) секреция билирубина из эндоплазматического ретикулума в желчь. Рассмотрим каждый из процессов в отдельности.
Поглощение билирубина печенью
Билирубин слаборастворим в плазме и воде; в плазме он специфически связывается с альбумином. Каждая молекула альбумина имеет, по-видимому, два центра связывания билирубина — высоко- и низкоаффинный. В 100 мл плазмы может содержаться 25 мг билирубина, прочно связанного с альбумином по его высокоаффинному центру. «Избыточный» билирубин связывается с альбумином менее прочно; он легко отделяется от альбумина, диффундируя в ткани. Ряд соединений — антибиотики и некоторые другие лекарственные вещества — конкурируют с билирубином за высокоаффинный центр альбумина. Эти соединения могут вытеснять билирубин из комплекса с альбумином и проявляют значительное клиническое действие.
В печени происходит переход билирубина от альбумина на синусоидальную поверхность гепатоцитов при участии насыщаемой системы переноса, в функционировании которой участвует некий переносчик. Эта система облегченного транспорта имеет очень большую емкость и даже при патологических условиях не лимитирует скорость метаболизма билирубина.
Поскольку система облегченного транспорта обеспечивает установление равновесия билирубина по обе стороны синусоидальной мембраны гепатоцита, поглощение билирубина зависит от его потребления в последующих метаболических процессах.
Конъюгация билирубина
В печени к билирубину присоединяются полярные группы и он переходит в водорастворимую форму, которая секретируется в желчь. Процесс, обеспечивающий повышение растворимости в воде (т. е. повышение полярности) билирубина, называется конъюгацией. Этот процесс, по крайней мере на начальных стадиях, протекает в гладком эндоплазматическом ретикулуме и осуществляется специальным набором ферментов. У млекопитающих билирубин секретируется в желчь преимущественно в форме билирубиндиглюкуронида (рис. 33.13). Сначала происходит образование билирубинмоноглюкуронида, которое катализируется UDP-глюкуронилтрансферазой — ферментом, присутствующим в гладком эндоплазматическом ретикулуме и состоящим, вероятно, из нескольких компонентов. Катализируемая реакция представлена на рис. 33.14. Она протекает главным образом в печени, а также в почках и слизистой кишечника. При нарушении метаболизма билирубина его конъюгаты находятся в сыворотке преимущественно в форме моноглюкуронидов.
Образование диглюкуронида билирубина может происходить в канальцах мембраны гепатоцитов при участии UDP-глюкуронилтрансферазы, подобной рассмотренной выше (рис. 33.14) или другого фермента дисмутазы, которая катализирует превращение двух молекул билирубинмоноглюкуронида в молекулу билирубиндиглюкуронида и молекулу свободного билирубина (рис. 33.14). Система конъюгации билирубина будет рассматриваться ниже в связи с наследственными нарушениями ее работы.
Активность UDP-глюкуронилтрансферазы может индуцироваться рядом используемых в клинике лекарств, в частности фенобарбиталом.
Секреция билирубина в желчь
Секреция конъюгированного билирубина в желчь идет против весьма высокого градиента концентрации и должна осуществляться с помощью механизма активного транспорта. Активный транспорт является, вероятно, скорость-лимитирующей стадией всего процесса метаболизма билирубина в печени. Транспорт конъюгированного билирубина из печени в желчь индуцируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, системы конъюгации билирубина и его вывода из гепатоцитов работают как единый функционально коррдинируемый механизм.

Рис. 33.13. Структура билирубиндиклюкуронида (конъюгированный «прямой» билирубин). Глюкуроновая кислота присоединяется эфирной связью к двум группам пропионовой кислоты с образованием ацилглюкуронида.
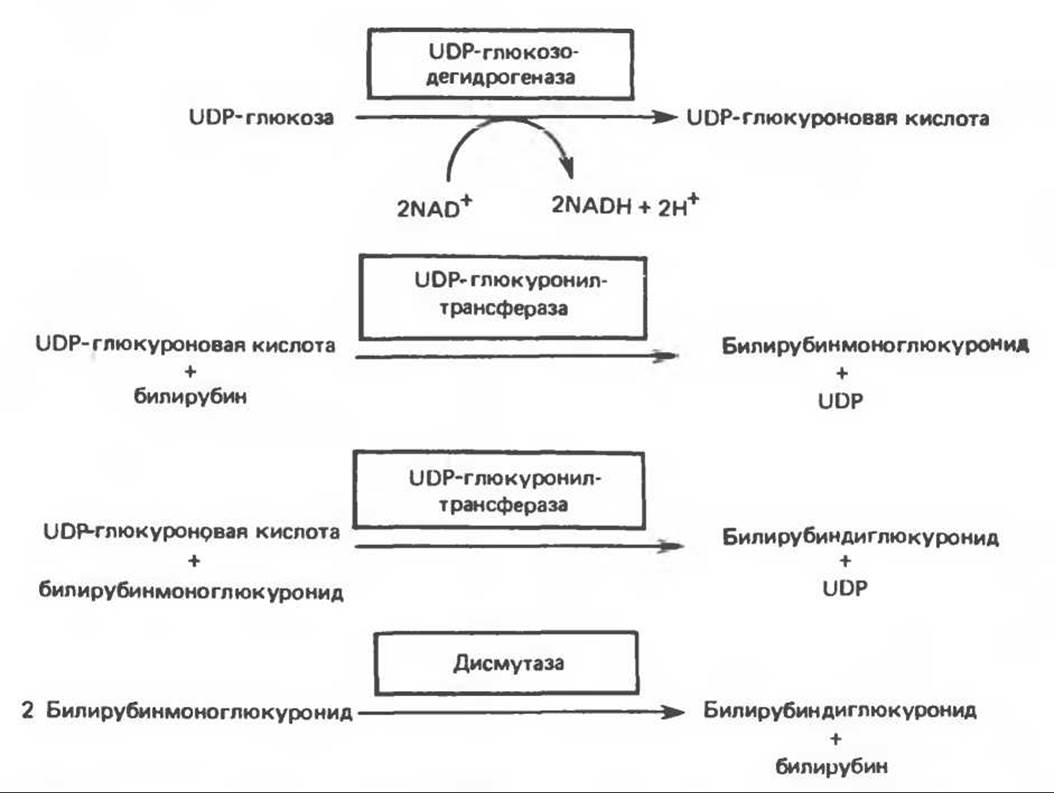
Рис. 33.14. Конъюгирование билирубина с глюкуроновой кислотой. Донором глюкуроната является UDP-глюкуроновая кислота, образующаяся из UDP-глюкозы.
При физиологических условиях практически весь секретируемый в желчь билирубин (свыше 97%) находится в конъюгированной форме. Только после светотерапии заметные количества неконъюгированного билирубина могут быть обнаружены в желчи.
В печени имеются многочисленные системы секреции в желчь природных и синтетических лекарственных соединений после их метаболизма. Некоторые из этих систем используются также билирубиндиглюкуронидами.