Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Гормоны половых желез
Биосинтез и метаболизм гормонов семенников
Синтез
Андрогены семенников синтезируются клетками Лейдига в интерстициальной ткани: в этих клетках содержится практически вся 3β-гидроксистероид-дегидрогеназа семенников — фермент, катализирующий ключевой этап биосинтеза тестостерона.
А. Биосинтетические пути.
1. Тестостерон служит непосредственным предшественником половых стероидов, подобно тому, как холестерол служит предшественником кортикостероидов надпочечников. Скорость-лимитирующим этапом, как и в надпочечниках, является отщепление боковой цепи холестерола. Превращение холестерола в прегненолон в надпочечниках, яичниках и семенниках происходит идентично. Однако в двух последних тканях реакция стимулируется не АКТГ, а ЛГ.
Превращение прегненолона в тестостерон протекает с участием пяти ферментов: 1) 3ß-гидроксистероид-дегидрогеназы (3βОН-СД); 2) ∆5,4-изомеразы; 3) 17а-гидроксилазы; 4) С17-20-лиазы и 5) 17β-гидроксистероид-дегидрогеназы (17β-ОН-СД). Соответствующая последовательность реакций, получившая название прогестеронового (или ∆4) пути, показана на рис. 50.1 справа. Превращение прегненолона в тестостерон может происходить также и по дегидроэпиандростероновому (или ∆5) пути (рис. 50.1, слева). В семенниках человека, по-видимому, преобладает ∆4-путь. Важно помнить, однако, что семенники человека мало доступны для исследования и большинство работ по выявлению этих путей проводилось на животных. Возможны существенные видовые различия.
Перечисленные пять ферментов локализованы в микросомной фракции семенников крысы, причем обнаружена тесная функциональная связь между активностями 3β-ОН-СД и ∆5,4-изомеразы и между активностями 17а-гидроксилазы и С17-20-лиазы. Эти ферментные пары изображены в общей последовательности реакции на рис. 50.1 ив схеме путей биосинтеза андрогенов в микросомных мембранах тестикулов на рис. 50.2. На последнем рисунке отражено поступление различных субстратов биосинтеза тестостерона в микросомы, а также последовательность их включения в цепь реакций ∆4-пути. В силу наличия четырех потенциальных субстратов для единственной, по-видимому, 3β-ОН-СД существует множество альтернативных путей. Путь, избираемый в каждом конкретном случае зависит, вероятно, от концентрации субстратов вблизи различных ферментов. Изменения концентраций могут быть обусловлены разделением субстратов в микросомной мембране.
2. Другие гормоны семенников. Дигидротестостерон (ДГТ) образуется из тестостерона в результате восстановления кольца А под действием фермента 5а-редуктазы. Суточная секреция ДГТ семенниками человека составляет примерно 50—100 мкг, однако подавляющая часть ДГТ является продуктом периферического превращения (см. ниже).
В семенниках вырабатываются также небольшие, но все же существенные количества 17β-эстрадиола (Е2) — женского полового гормона. Большая часть образующегося у самцов Е2 — это результат периферической ароматизации тестостерона и андростендиона. Считается, что в синтезе Е2 участвуют клетки Лейдига, клетки Сертоли и семенные канальцы. Роль Е2 у самцов не установлена. Возможно, он участвует в механизмах регуляции ФСГ. Чрезмерно высокое содержание Е2 в плазме и изменения соотношения свободный Е2: тестостерон характерны для пубертатной или постпубертатной гинекомастии, а также для хронических болезней печени или при гипертиреозе.
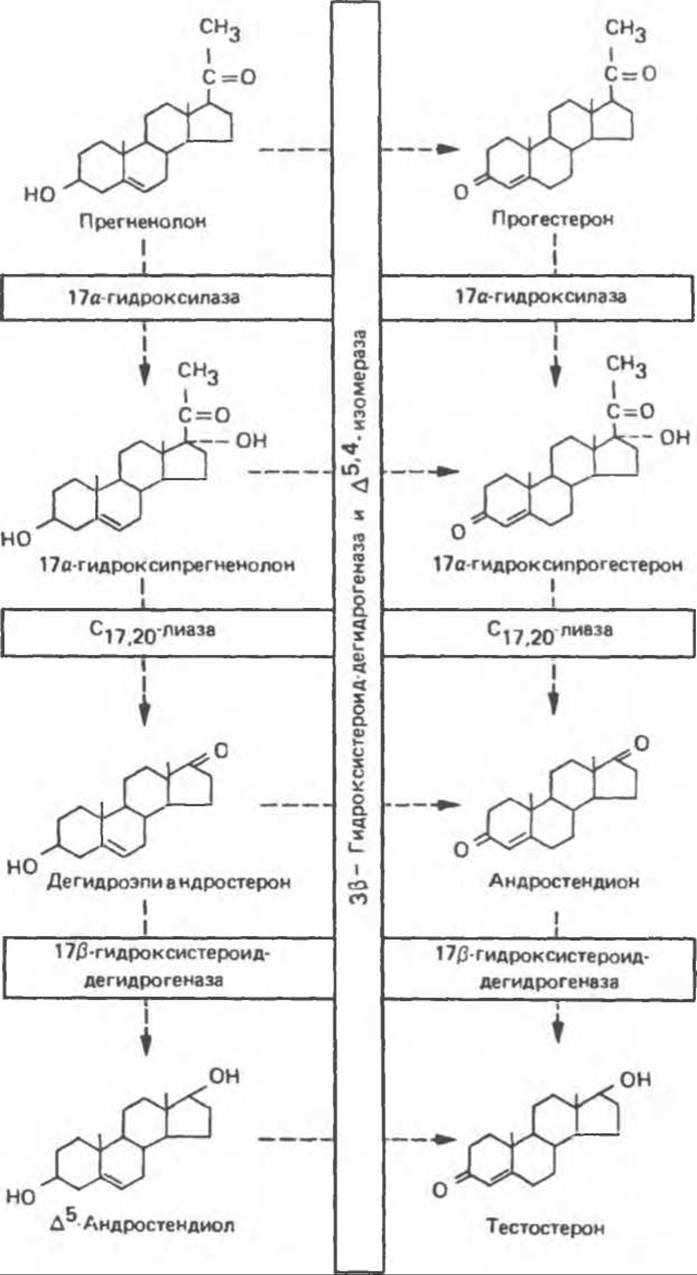
Рис. 50.1. Пути биосинтеза тестостерона. Слева дегидроэпиандростероновый (или ∆5) путь, справа прогестероновый (или ∆4) путь.
Б. Возрастные изменения продукции тестикулярных гормонов
У плодов и новорожденных крыс преобладает тестостерон. Однако вскоре после рождения семенники начинают производить только андростерон. Способность к образованию тестостерона восстанавливается в период полового созревания и сохраняется до конца жизни. Аналогичные результаты получены и в отношении других видов. Вполне возможно, что такие же возрастные изменения наблюдаются у человека.
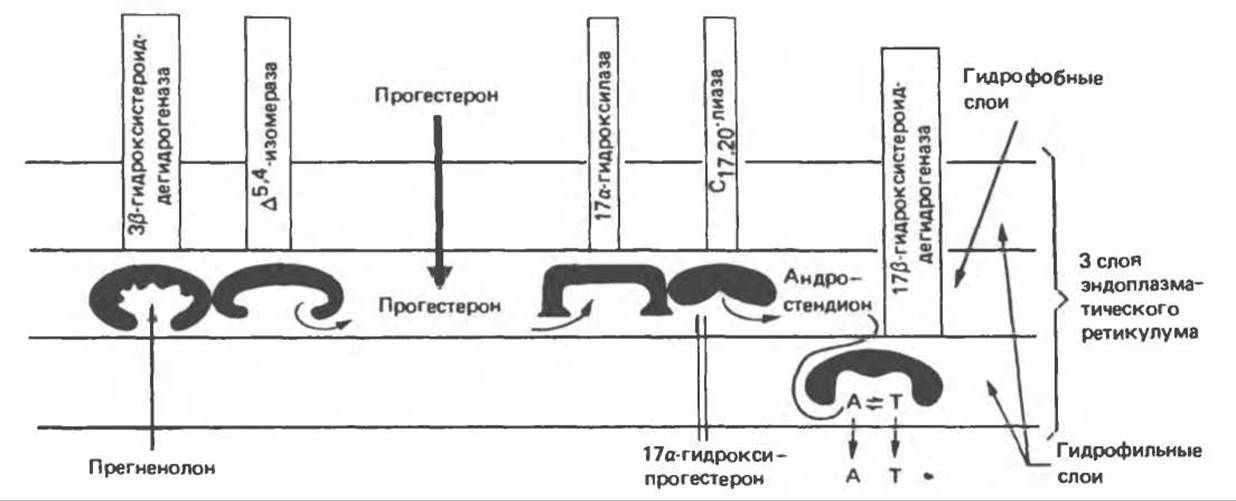
Рис. 50.2. Схема биосинтеза андрогенов в микросомной мембране семенников. Мембрана изображена в виде горизонтальной линии, что, возможно, соответствует ее виду в клетке. Однако в препаратах микросом она образует пузырьки (везикулы). А — андростендион, Т — тестостерон. (Reproduced, with permission, from DeGroot L. J. Endocrinology, Vol. 3, Grune and Stratton, 1979.)
Секреция и транспорт
В венозной крови семенников присутствует несколько стероидов, но главный стероид, секретируемый семенниками взрослой особи, — это тестостерон. У мужчин суточная секреция тестостерона составляет в норме 5 мг. Процесс секреции тестикулярных стероидов, по-видимому, не регулируется; как и другие стероидные гормоны, тестостерон, очевидно, секретируется по мере образования.
Таблица 50.1. Связывание гормонов с секс-гормон-связывающим глобулином (СГСГ)
|
Связываемые стероиды |
Несвязываемые стероиды |
|
Тестостерон |
Конъюгированные андрогены |
|
n-ß-эстрадиол |
17-а-тестостерон |
|
Дигидротестостерон |
Дегидроизоандростерон |
|
Другие n-ß-гидрокси-стероиды |
Кортизол |
|
Эстрон |
Прогестерон |
В плазме большинства млекопитающих, в том числе и человека, имеется ß-глобулин, специфически связывающий тестостерон с относительно высоким сродством и ограниченной емкостью (табл. 50.1). Этот белок, называемый обычно секс-гормон-связывающим глобулином (СГСГ) или тестостерон-эстроген-связывающим глобулином (ТЭСГ), образуется в печени. Его продукция усиливается под действием эстрогенов (у женщин концентрация СГСГ в сыворотке вдвое выше, чем у мужчин) при определенных болезнях печени и при гипертиреозе; под влиянием андрогенов с возрастом и при гипотиреозе образование данного белка уменьшается. Многие из этих факторов влияют также на синтез кортикостероид-связывающего глобулина (см. гл. 48) и тиреотропин-связывающего глобулина (см. гл. 46). Поскольку СГСГ и альбумин связывают до 97—99% циркулирующего в крови тестостерона, лишь небольшая его часть находится в крови в свободной (биологически активной) форме. Основная функция СГСГ состоит, вероятно, в том, чтобы ограничивать концентрацию свободного тестостерона в сыворотке. Тестостерон связывается с СГСГ с большим сродством, чем эстрадиол (табл. 50.2), поэтому при изменении концентрации СГСГ содержание свободного тестостерона меняется в большей степени, чем свободного эстрадиола. Увеличение концентрации СГСГ способствует росту соотношения свободный.
Таблица 50.2. Приблизительное сродство стероидов к связывающим белкам плазмы эстрадиолгтестостерон. Этот феномен имеет место при старении, циррозе печени и гипертиреозе и, следовательно, вносит определенный вклад в признаки и симптомы «эстрогенизации», свойственной этим состояниям.
|
Гормон |
СГСГ 1) |
КСГ 1) |
|
Эстрадиол |
5 |
> 10 |
|
Эстрон |
> 10 |
> 100 |
|
Андростендион |
... |
... |
|
Тестостерон |
2 |
> 100 |
|
Дигидротестостерон |
1 |
> 100 |
|
Прогестерон |
> 100 |
2 |
|
Кортизол |
> 100 |
3 |
1) Сродство выражается как в молях х 109. (Adapted from Siiteri R. К., Febres F. Ovarian hormone synthesis, circulation and mechanisms of action. Page 1401. In: Endocrinology, Vol. 3. DeCroot L. J. (editor). Grune and Stratton, 1979.)
Периферический метаболизм и экскреция
А. Метаболические пути. Метаболические превращения тестостерона осуществляются двумя путями. Один путь включает в себя окисление в 17-м положении, другой — восстановление двойной связи кольца А и 3-кетогруппы1. В результате первого пути, функционирующего во многих тканях, в том числе и в печени, образуются 17-кетостероиды как правило, лишенные активности или обладающие более слабой активностью, чем исходное соединение. Второй путь, менее эффективный, протекает главным образом в тканях-мишенях и ведет к образованию активного метаболита — ДГТ, а также эстрадиола и андростандиола. Этиохоланолон и андростерон — это 5 ß-восстановленные продукты андрогенов.
Б. Метаболиты тестостерона. Наиболее важный метаболит тестостерона — ДГТ — представляет собой активную форму гормона и обнаруживается во многих тканях, включая семенные пузырьки, предстательную железу, наружные половые органы и некоторые участки кожи. В плазме взрослых мужчин содержание ДГТ примерно в десять раз ниже содержания тестостерона: за сутки его образуется приблизительно 400 мкг (сравните с 5 мг тестостерона). Превращение тестостерона в ДГТ катализируется NADPH-зависимой 5а-редуктазой.
Таким образом, тестостерон можно рассматривать как прогормон по двум причинам: во-первых, он превращается в более активное соединение дигидротестостерон и, во-вторых, превращение это происходит главным образом в тканях, расположенных вне семенников. Небольшая часть тестостерона ароматизируется, образуя эстрадиол, что особенно важно для мозга, где эти гормоны участвуют в формировании полового поведения животных. Андростандиол, еще один высоко активный андроген, также образуется из тестостерона.
1 По новой номенклатуре следует писать оксогруппа, оксостероиды и т. д., но поскольку в медицинской литературе принят термин «кетостероиды», здесь мы будем придерживаться этого названия.— Прим. перев.
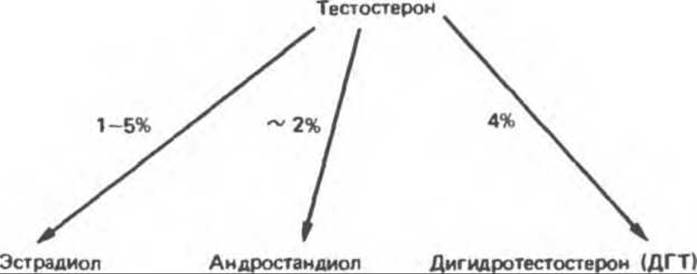
Главные 17-кетостероидные метаболиты тестостерона — андростерон и этиохоланолон — конъюгируют в печени с глюкуронидом и сульфатом с образованием водорастворимых экскретируемых соединений. Количественное определение 17-кетостероидов в моче использовалось ранее в качестве теста на андрогенную активность. Теперь, однако, установлено, что этот показатель слабо отражает гормональный статус in vivo.