Биохимия человека Том 2 - Марри Р. 1993
Структура, функция и репликация информационных макромолекул
Технология рекомбинантных ДНК
Некоторые практические приложения технологии рекомбинантной ДНК
Выделение специфического гена из целого генома требует методики, с помощью которой можно среди миллиона сходных элементов найти один, нужный исследователю. Идентификация регуляторной последовательности длиной в 10 нуклеотидов требует чувствительности, соответствующей выявлению 1 элемента из 3 х 108. Серповидноклеточная анемия вызвана заменой только 1 основания в геноме, т. е. 1 элемента из 3 х 109. Достигнутый сегодня уровень развития методов генной инженерии достаточен для работы на таком уровне чувствительности.
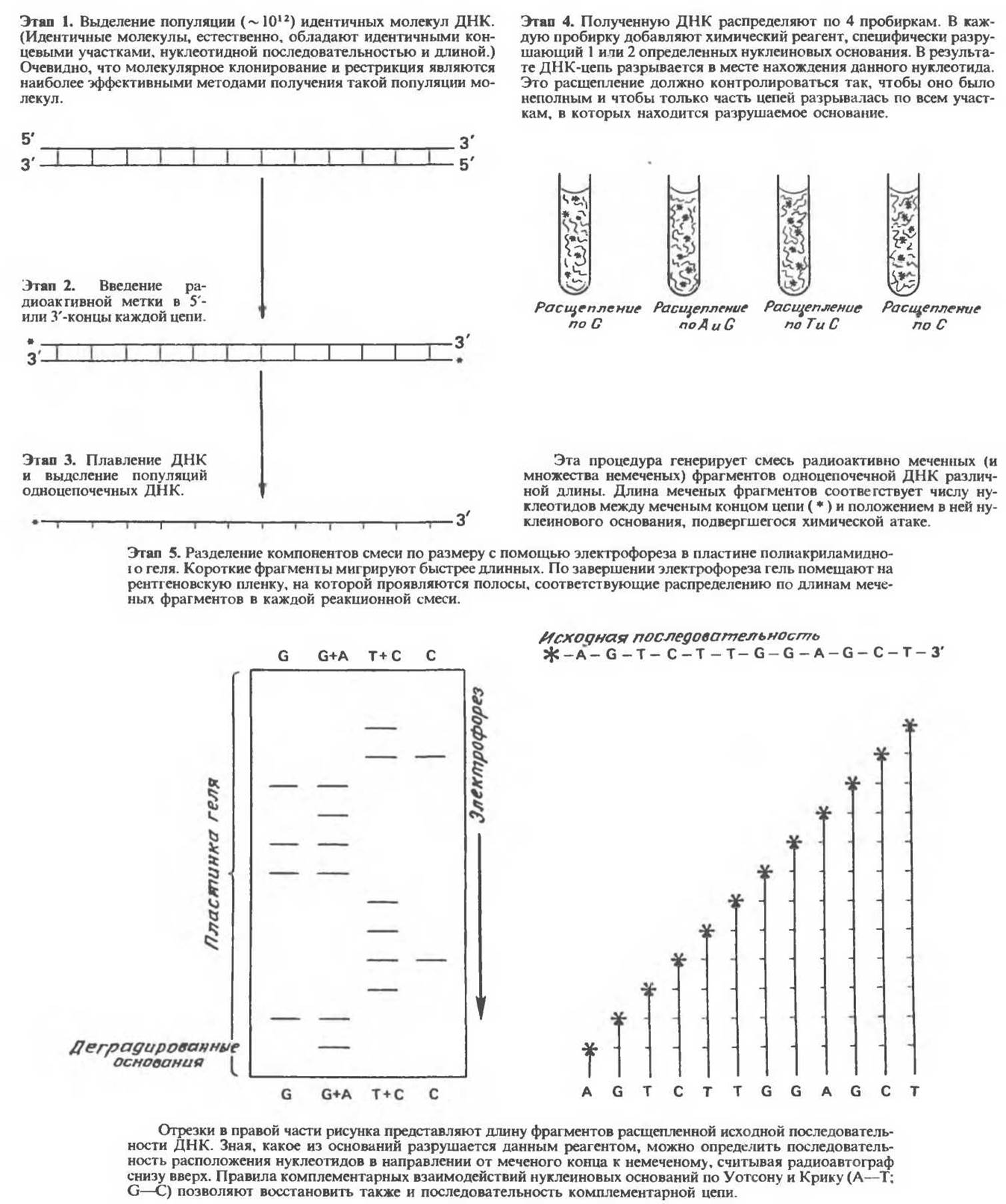
Рис. 36.6. Определение нуклеотидной последовательности ДНК (секвенирование ДНК) по Максаму—Гилберту.
Генетическое картирование
Под этим термином подразумевается совокупность подходов и методов, с помощью которых можно каждый ген отнести к определенной хромосоме, т. е. составить генетическую карту организма. Например, у человека благодаря применению двух основных методов — гибридизации соматических клеток и гибридизации in situ — установлена хромосомная локализация ряда генов, ответственных за некоторые заболевания. При гибридизации in situ препарат метафазных хромосом на поверхности стеклянной пластины инкубируют с радиоактивно меченным зондом. Точную область гибридизации определяют с помощью радиоавтографии (фотографическую эмульсию наносят, непосредственно на пластинку). Образование зерен над гистологически идентифицированной хромосомой позволяет сделать вывод о принадлежности данного гена к конкретной хромосоме, а часто и к определенному ее участку. Некоторые гены человека, локализованные методом гибридизации in situ, представлены в табл. 36.5.
Нет сомнения, что в ближайшие годы карта генома человека станет полной: более того, обсуждается принципиальная возможность определения полной нуклеотидной последовательности человеческого генома. Однако уже сейчас, опираясь на имеющиеся данные, можно сделать ряд существенных заключений.
1. Гены, кодирующие белки со сходными функциями, могут находиться в разных хромосомах (а- и ß-глобины).
2. Гены, относящиеся к одному семейству, также могут локализоваться в разных хромосомах (гормон роста и пролактин).
3. Гены, детерминирующие многие наследственные патологии, вызванные недостаточностью специфических белков (в том числе сцепленные с X- хромосомой), действительно расположены в совершенно определенных сайтах хромосом.
Таблица 36.5. Локализация генов человека
|
Ген |
Хромосома |
Заболевание |
|
Инсулин |
11р15 |
|
|
Пролактин |
6p23-q12 |
|
|
Гормон роста |
17q21-qter |
Недостаточность гормона роста |
|
а-Глобин |
16p12-pter |
а-Талассемия |
|
ß-Глобин |
11p12 |
ß-Талассемия, серповидные клетки |
|
Аденозин-дезаминаза |
20q13-qter |
Недостаточность аденозин-дезаминазы |
|
Фенилаланин-гидроксилаза |
12q24 |
Фенилкетонурия |
|
Гипоксантин-гуанин-фосфорибозилтрансфераза |
Xq26-q27 |
Синдром Леша—Найхана |
|
Сегмент ДНК G8 |
4p |
Хорея Гентингтона |
В таблице представлены хромосомные локализации нескольких генов и болезни, вызванные нарушениями в продуктах этих генов. Первой (выделенной) цифрой или буквой указана хромосома. Остальные цифры и буквы детализируют внутрихромосомную локализацию по McKusick V. A. «Mendelian Inheritance in Man» 6th ed. Johns Hopkins Univ. Press. 1983.
Благодаря использованию клонированных фрагментов установлена хромосомная локализация многих генетических нарушений, для которых не удавалось выявить недостаточности по каким-либо специфическим белкам. К таким заболеваниям относятся: хорея Гентингтона (хромосома 4); муковисцидоз (хромосома 7); поликистозная нефропатия взрослых (хромосома 16); мышечная дистрофия Дюшенна (X-хромосома). Если область ДНК, в которой локализован дефект, имеет характерную структуру гена (рис. 36.1), то можно синтезировать этот ген, ввести в соответствующий вектор, добиться экспрессии и изучать функцию. Кроме того, можно синтезировать олигопептид, последовательность аминокислот в котором определяется согласно установленной открытой рамке считывания в кодирующей области. Антитела, полученные против этого пептида, представляют собой инструмент для выявления экспрессии данного пептида (или констатации ее отсутствия) у здоровых и больных людей.
Получение белков
Одно из практических применений технологии рекомбинантных ДНК — получение медикобиологической продукции. Генная инженерия дает возможность получать в больших количествах белки, которые не могут быть выделены применением обычных методов очистки (интерферон, плазминоген-активирующий фактор); кроме того, с помощью рекомбинантных ДНК можно нарабатывать специфические белки человека для замены используемых в клинической практике аналогичных белков животных (инсулин, гормон роста). Достоинства обеих технологий очевидны.
Первоначально цели генной инженерии ограничивались получением веществ (как правило, белков) для лечения (инсулин), диагностики (тест на СПИД) и профилактики (вакцина против вируса гепатита В) болезней человека. В настоящее время представления о возможностях биотехнологии значительно расширились. Так, уже осуществляются попытки сконструировать растения, устойчивые к засухе и экстремальным температурам, а также более эффективно фиксирующие азот.

Рис. 36.7. Схема кластера генов ß-глобина и некоторые генетические нарушения, приводящие к заболеваниям. Ген ß- глобина расположен в 11-й хромосоме в непосредственной близости от двух генов у-глобинов и гена 6-глобина. Семейство ß-генов организовано в последовательность 5'-ξ-Gy-Ay-ψß-δ-ß-3'. Локус ξ экспрессируется на ранних этапах жизни эмбриона (а2ξ2). Гены у экспрессируются на стадии плода, образуя фетальный гемоглобин (HbF, а2у2). Г емоглобин взрослых состоит из HbA (a2ß2) или НbА2(а2δ2). Ген ψß является псевдогеном; его последовательность гомологична последовательности ß-гена, но включает мутации, препятствующие экспрессии. Делеции (затемненные участки) ß-локуса вызывают ß-талассемию (недостаточность или отсутствие [ß°] ß-глобина). Делеции δ- и ß-генов приводят к образованию гемоглобина Лепоре (образуется только а-гемоглобин). Инверсия (Ayδβ) этой области (незатемненный участок) полностью ингибирует функцию гена и обусловливает талассемию (тип III). Каждый тип талассемии характерен для определенных этнических групп. Так (Аyδβ)°-талассемия встречается в основном у выходцев из Индии. В этой области генома картированы и многие другие делеции, вызывающие определенный тип талассемии.