Биохимия человека Том 2 - Марри Р. 1993
Структура, функция и репликация информационных макромолекул
Синтез белка и генетический код
Транспортные РНК и синтез белка
Для каждой из 20 аминокислот существует по крайней мере один вид тРНК. Все молекулы тРНК характеризуются необычайным сходством как в функциональном отношении, так и по пространственной структуре. Выполнение адапторных функций становится возможным после образования специфического комплекса между молекулой тРНК и определенной аминокислотой. Так как у нуклеиновых кислот нет какого-либо специального сродства к боковым цепям аминокислот, взаимное узнавание должно происходить с помощью специальной молекулы белка, способной выявлять одновременно и определенную тРНК-молекулу, и соответствующую аминокислоту. Для подобного узнавания и правильного присоединения соответствующей аминокислоты к молекуле тРНК должно существовать по крайней мере 20 специфичных ферментов. Процесс узнавания и присоединения происходит в два этапа и катализируется ферментом — уникальным для каждой из 20 аминокислот, принадлежащим к классу аминоацил-тРНК-синтетаз. Этот фермент образует активированный промежуточный аминоацил-АМР-ферментный комплекс (рис. 40.1), который специфически узнает соответствующую молекулу тРНК и переносит аминокислотный остаток на 3'-гидроксильную группу концевого аденозина. Аминокислота остается присоединенной эфирной связью к тРНК вплоть до включения в определенное положение растущей полипептидной цепи предшественника белка.
Теперь вернемся к структуре молекулы тРНК, описанной в гл. 37 и изображенной на рис. 37.11. Тимидин-псевдоуридин-цитидиновое плечо (ТΨС) участвует в связывании аминоацил-тРНК с рибосомой в сайте синтеза белка. Плечо D необходимо для специфического узнавания данной молекулы тРНК соответствующей аминоацил-тРНК-синтетазой. Акцепторное плечо служит местом присоединения соответствующей аминокислоты.
Антикодоновый участок состоит из 7 нуклеотидов и узнает трехбуквенный кодон в мРНК (рис. 40.2). Последовательность этой области, если читать ее от 3'- к 5'-концу, имеет следующую структуру: варьирующее основание, модифицированный пурин, собственно антикодон (х, у, z), пиримидин, пиримидин- 5'. Обратите внимание, что антикодон считывается в направлении 3'→5', а генетический код от 5' к 3', так как кодон в мРНК и антикодоновая петля в гРНК — антипараллельны.
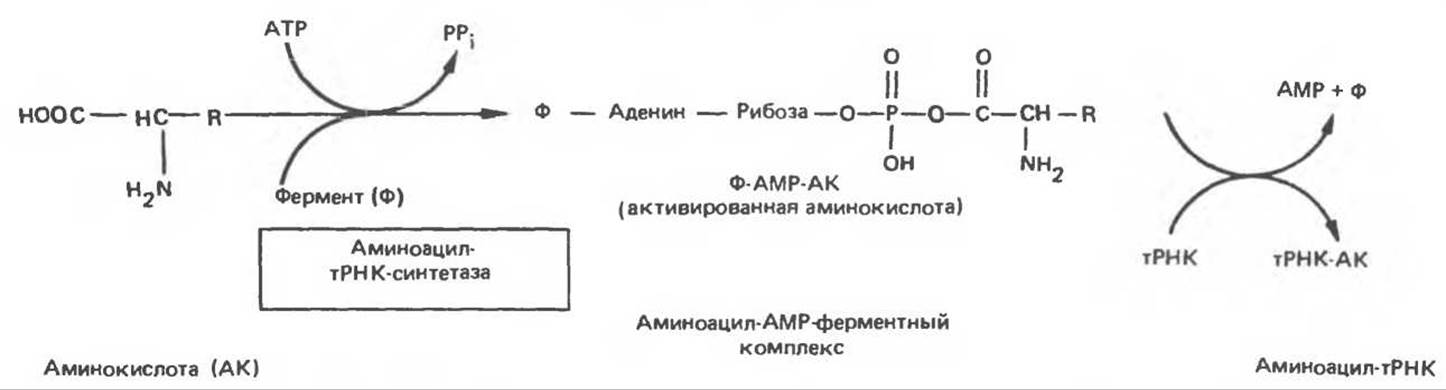
Рис. 40.1. Образование аминоацил-тРНК. Двухступенчатая реакция с участием аминоацил-тРНК-синтетазы приводит к образованию аминоацил-тРНК. В первой реакции образуется комплекс АМР—аминокислота фермент. Активированная аминокислота переносится на соответствующую тРНК-молскулу. АМР и фермент высвобождаются, после чего фермент может снова участвовать в синтетазной реакции.
Вырожденность генетического кода касается в основном третьего нуклеотида кодона и предполагает, что образование комплементарной пары между ним и соответствующим нуклеотидом антикодона не должно быть абсолютно строгим. Как уже упоминалось, это явление принято называть неполным соответствием или качанием, поскольку в области взаимодействия последнего нуклеотида кодона с антикодоном допускается нестрогое связывание — «качание». Например, 2 кодона аргинина AGA и AGG могут специфически связываться с одним и тем же антикодоном, содержащим на 5'-конце остаток урацила. Аналогично три кодона глицина GGU, GGC и GGA могут образовывать пары с антикодоном CCI. Инозин (I) — это еще одно из необычных (минорных) оснований, встречающихся в структуре тРНК.
Узнавание кодона молекулой тРНК не зависит от того, какая аминокислота присоединена к ее 3'-концу. Это было продемонстрировано в эксперименте, в котором радиоактивный цистеин, присоединенный к специфической молекуле тРНК (tPHKcvs), химическим путем превращали в аланин. При этом антикодоновый участок самой тРНКсуs оставался неизменным. При использовании такой аланил-тРНКсys в трансляции гемоглобиновой мРНК радиоактивный аланин обнаруживали в аминокислотной последовательности в положениях, в норме занимаемых остатками цистеина. Этот эксперимент продемонстрировал, что сам аминокислотный остаток не принимает участия в специфическом узнавании кодона. Как уже отмечалось, встраиваемые в полипептидную цепь аминокислотные остатки вообще не контактируют непосредственно с мРНК-матрицей.
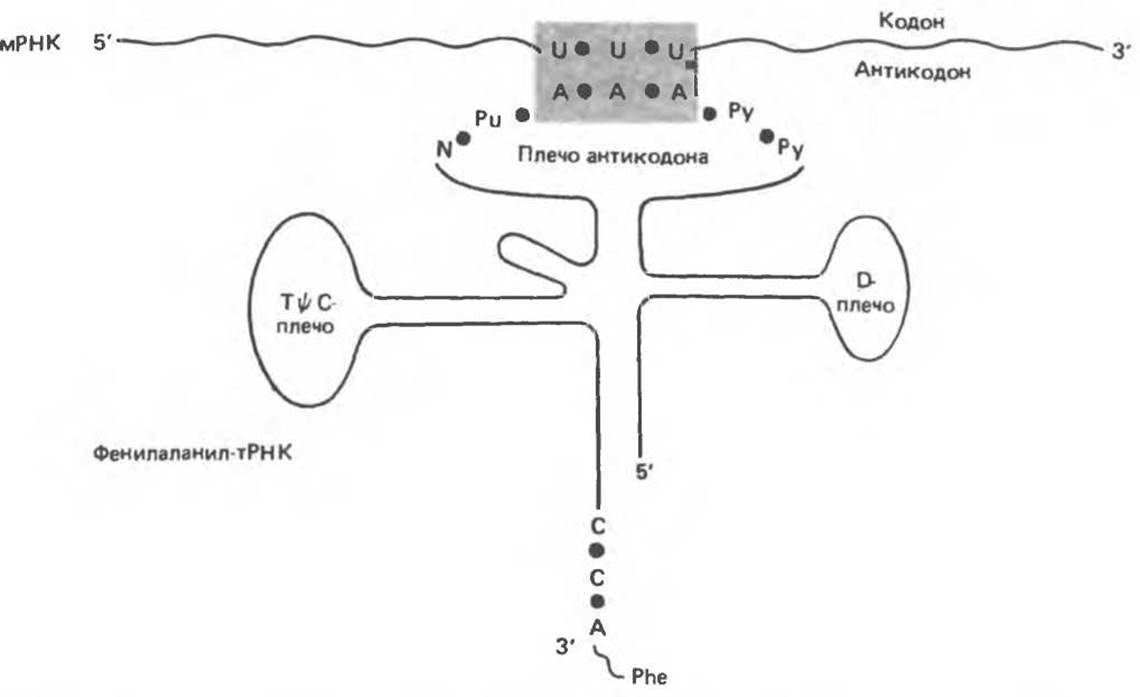
Рис. 40.2. Узнавание кодона антикодоном. UUU — один из кодонов фенилаланина. Молекула тРНК, «заряженная» фенилаланином (Phe), содержит в антикодоновом участке комплементарную последовательность ААА, образующую комплекс с кодоном из трех пар оснований. Антикодоновый участок, как правило, содержит следующую гептануклеотидную последовательность: варьируемый нуклеотид (N); модифицированный пурин (Pu*); антикодон (X, Y, Z): два пиримидина (Ру) (ориентация 3'→5').