Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Мембраны: структура, сборка и функции
Трансмембранный перенос малых молекул
Молекулы могут пассивно пересекать бислой по электрохимическому градиенту путем простой или облегченной диффузии. Такому спонтанному переносу, приводящему к установлению равновесия, противостоит активный транспорт, который требует затрат энергии, поскольку он происходит против электрохимического градиента. Эти механизмы схематически представлены на рис. 42.13.
Пассивная диффузия
Как мы уже говорили, некоторые вещества, например газы, могут проникать в клетку за счет трансмембранной диффузии по электрохимическому градиенту; при этом никаких энергетических затрат не требуется. Скорость простой диффузии через мембрану растворенных веществ определяется тепловым движением перемещающихся молекул, трансмембранным концентрационным градиентом вещества и его растворимостью (коэффициентом проницаемости; рис. 42.6) в гидрофобном слое мембраны. Растворимость обратно пропорциональна числу водородных связей, которые должны быть разорваны, чтобы растворенное в водной среде вещество оказалось включенным в гидрофобный слой. Электролиты, слабо растворимые в липидах, не образуют с водой водородных связей, но они обладают водной оболочкой, образующейся в результате электростатических взаимодействий. Размер оболочки прямо пропорционален плотности заряда электролита. Электролиты с большей плотностью заряда обладают большей гидратной оболочкой и, таким образом, меньшей скоростью диффузии. Ионы Na+, например, характеризуются большей плотностью заряда, чем ионы К+. Следовательно, гидратированный Na+ имеет больший размер, чем К+, и его скорость пассивной диффузии ниже.
В природных мембранах в отличие от синтетических бислойных мембран имеются трансмембранные каналы — сходные с порами структуры, состоящие из белков. Каналы, пропускающие катионы, имеют средний диаметр ~ 5—8 нм и выстланы отрицательно заряженными группами. Проводимость канала зависит от размера, степени гидратации и плотности заряда иона. Обнаружены специальные каналы для Na+, К+ и Са2+.
В мембранах нервных клеток имеются хорошо изученные ионные каналы, ответственные за генерацию и распространение потенциала действия вдоль мембраны. Активность некоторых из них контролируется нейромедиаторами, т.е. работа каналов может регулироваться. Кроме того, один ион может регулировать активность канала, проницаемого для другого иона. Так, при уменьшении концентрации Са2+ во внеклеточной жидкости увеличивается мембранная проницаемость и диффузия Na+. В результате мембрана деполяризуется и генерируется нервный импульс. Именно этим объясняются оцепенелость, покалывание и судороги мышц при понижении уровня Са2+ в плазме.
Таблица 42.4. Перенос вещества и информации через мембраны
|
Трансмембранное перемещение малых молекул Диффузия (пассивная и облегченная) Активный транспорт |
|
Трансмембранное перемещение крупных молекул Эндоцитоз Экзоцитоз |
|
Передача сигнала через мембраны Рецепторы клеточной поверхности 1. Передача сигнала (например, глюкагон→сАМР) 2. Интернализация сигнала (сопряженная с эндоцитозом, например рецептор ЛНП) Движение внеклеточных рецепторов (стероидные гормоны; некая разновидность диффузии) |
|
Межклеточные контакты и коммуникации |
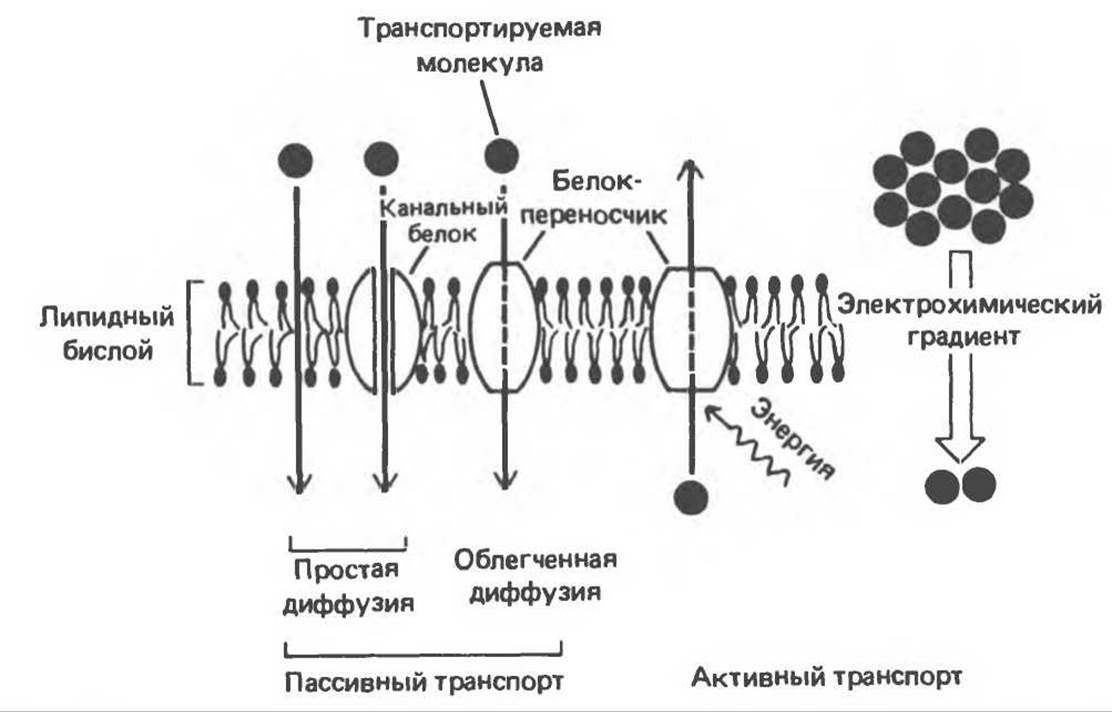
Рис. 42.13. Многие мелкие незаряженные молекулы свободно проходят через липидный бислой. Заряженные молекулы, крупные незаряженные молекулы и некоторые мелкие незаряженные молекулы проходят через мембраны по каналам или порам либо с помощью специфических белков-переносчиков. Пассивный транспорт всегда направлен по электрохимическому градиенту в сторону установления равновесия. Активный же транспорт осуществляется против электрохимического градиента и требует энергетических затрат. (Из работы Alberts В. et al.: Molecular Biology of the cell. Garland, 1983.)
Каналы открываются только на определенное время, т. е. обладают воротным механизмом. В случае воротного механизма, контролируемого лигандом, некая специфическая молекула связывается с рецептором и открывает «ворота». Каналы с потенциалзависимым воротным механизмом открываются (или закрываются) в ответ на изменение мембранного потенциала.
Некоторые микроорганизмы синтезируют малые органические молекулы — ионофоры, которые осуществляют челночные перемещения ионов через мембраны. Эти ионофоры содержат гидрофильные центры, которые связывают определенные ионы. По периферии центры окружены гидрофобными областями, что позволяет молекуле легко растворяться в мембране и диффундировать через нее. Существуют и другие ионофоры, подобные хорошо изученному полипептиду грамицидину, которые образуют каналы. Некоторые микробные токсины (например, дифтерийный токсин) и компоненты активированного сывороточного комплемента способны образовывать крупные поры в клеточных мембранах, через которые могут проходить макромолекулы.
Суммируя сказанное, можно сказать, что диффузия веществ определяется следующими факторами:
1) трансмембранным концентрационным градиентом веществ. Растворенные вещества перемещаются в сторону понижения концентрации; 2) трансмембранной разностью электрических потенциалов. Растворенные вещества движутся в сторону раствора с противоположным зарядом; 3) коэффициентом проницаемости мембраны для данного вещества; 4) градиентом гидростатического давления на мембране. При повышении давления будет увеличиваться скорость столкновений молекул и мембраны; 5) температурой. Чем выше температура, тем больше скорость частиц и, следовательно, частота столкновений между частицами и мембраной.
Облегченная диффузия и активный транспорт
Транспортные системы можно описывать исходя из числа переносимых молекул и направления перемещения (рис. 42.14) или в соответствии с тем, как осуществляется перенос — в сторону установления равновесия или против него. В системе уннпорта происходит перенос молекулы одного типа в обоих направлениях. В системах котранспорта перенос одного растворенного вещества сопровождается переносом (одновременным или последовательным) стехиометрического количества другого. В случае симпорта оба вещества перемещаются в одном направлении. Примерами таких систем являются перенос Н+/сахара и Na+/сахара (глюкоза, галактоза, ксилоза и арабиноза) в бактериях и Na+/аминокислот в клетках млекопитающих. В случае антипорта вещества переносятся в противоположных направлениях (например, Na+ в клетку, а Са2+ из клетки).
Молекулы, которые сами не могут пересекать липидный бислой, используют для этого белки-переносчики, с которыми они связываются. Такое перемещение может происходить двумя способами: путем облегченной диффузии или активного транспорта с помощью высокоспецифичных транспортных систем.
Облегченная диффузия и активный транспорт во многом сходны. Оба процесса, по-видимому, осуществляются при участии специальных белков-переносчиков и для обоих характерна специфичность к ионам, сахарам и аминокислотам. Об этом свидетельствуют результаты анализа тех последствий, к которым приводят мутации в бактериальных и животных клетках (включая некоторые мутации, вызывающие заболевания у человека). Облегченная диффузия и активный транспорт напоминают реакцию между ферментом и субстратом, однако они осуществляются без образования ковалентных связей. На это сходство указывают следующие моменты:
1) имеется специфический участок связывания для растворенного вещества; 2) процесс переноса характеризуется насыщением, т. е. существует некая максимальная скорость транспорта Vmах (рис. 42.15); 3) процесс характеризуется определенной константой связывания, так что система в целом имеет свою Км (рис. 42.15); 4) вещества, сходные по своей структуре с переносимым соединением, являются конкурентными ингибиторами и блокируют транспорт.
Основные различия между облегченной диффузией и активным транспортом состоят в следующем:
1) облегченная диффузия может осуществляться в обоих направлениях, тогда как активный транспорт — обычно лишь в одном; 2) активный транспорт всегда идет против электрического или химического градиента и требует энергетических затрат.
А. Облегченная диффузия. Некоторые вещества диффундируют через мембраны по электрохимическому градиенту быстрее, чем можно ожидать исходя из их размеров, заряда или коэффициента распределения. Эта облегченная диффузия отличается по своим свойствам от простой диффузии. Скорость облегченной диффузии, осуществляемой по унипортному механизму, выходит на плато, т. е. число участков связывания данного вещества ограничено. Многие системы облегченной диффузии стереоспецифичны, но, как и в случае простой диффузиии, перенос осуществляется без энергетических затрат.
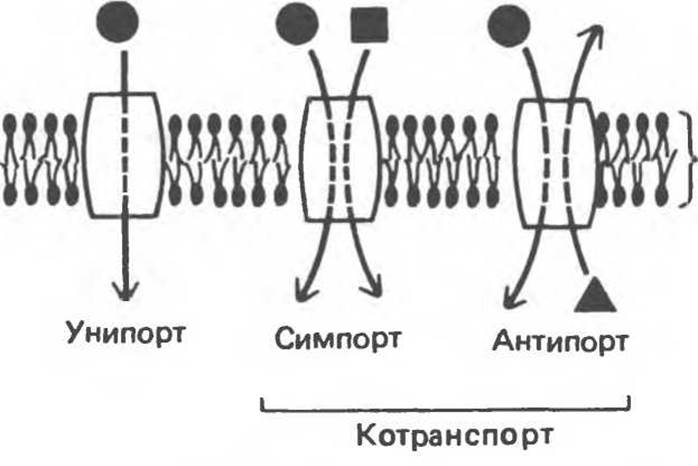
Рис. 42.14. Схематическое представление типов транспортных систем. Переносчики можно подразделить в соответствии с направлением перемещения вещества и степенью разнообразия переносимых молекул. (Из книги Alberts В. et al.: Molecular Biology of the cell. Garland, 1983.)
Как мы уже говорили, асимметричное распределение мембранных белков между внутренней и внешней сторонами мембраны достаточно стабильно, спонтанное перемещение белков через мембрану происходит исключительно редко, а, следовательно, в основе облегченной диффузии, по-видимому, не может лежать трансмембранное перемещение белков-переносчиков; исключение составляют ионофоры, присутствующие в мембранах бактериальных клеток.
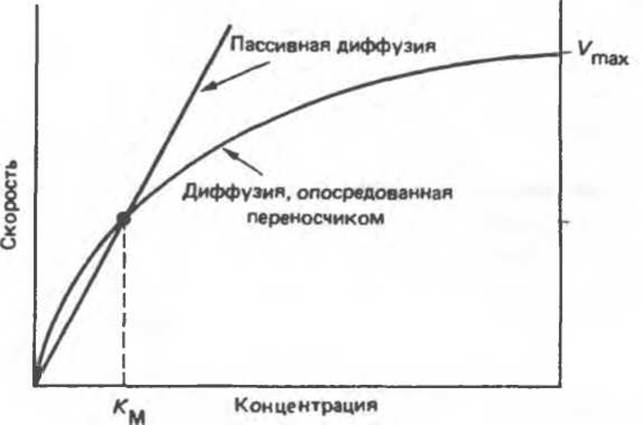
Рис. 42.15. Сравнение кинетик опосредованной переносчиком (облегченной) диффузии и простой диффузии. В последнем случае скорость перемещения вещества прямо пропорциональна его концентрации в растворе, тогда как при наличии переносчика наблюдается насыщение. Vmax — максимальная скорость. Константа Км равна такой концентрации вещества, при которой скорость составляет половину максимальной.
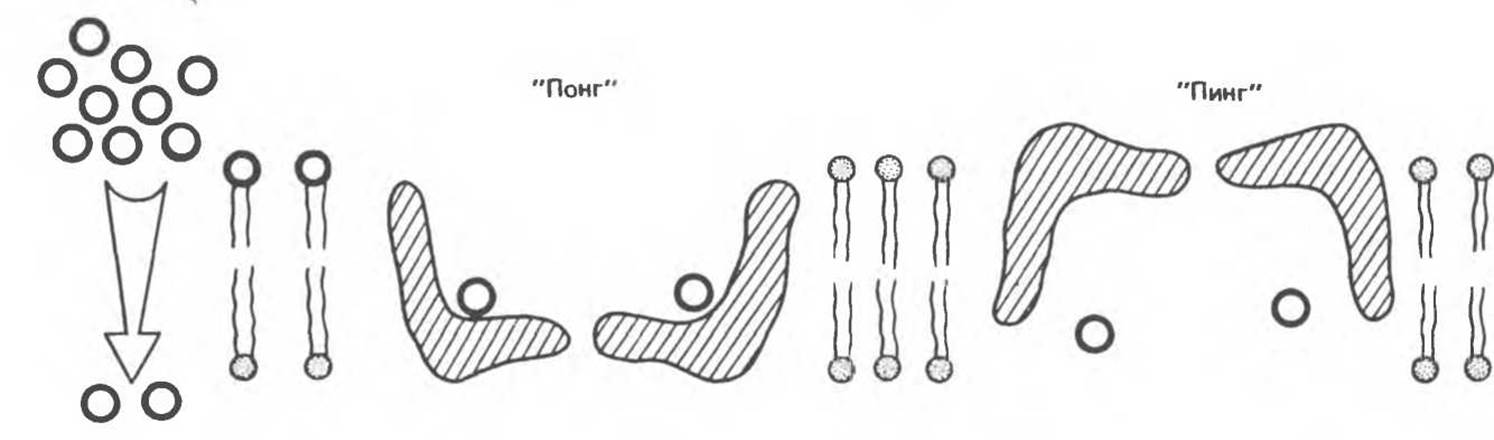
Рис. 42.16. Облегченная диффузия, механизм «пинг-понг». Белок-переносчик (заштрихован) связывает вещество, находящееся в растворе с высокой его концентрацией по одну сторону мембраны. Затем в переносчике происходят конформационные изменения («понг»→«пинг»), в результате которых это вещество высвобождается по другую сторону мембраны. Свободный переносчик возвращается в исходное состояние («пинг»→«понг»), и цикл завершается.
Процесс облегченной диффузии можно объяснить с помощью механизма «пинг-понг» (рис. 42.16). Согласно этой модели, белок-переносчик может находиться в двух основных конформациях. В состоянии «понг» он экспонирован в раствор с высокой концентрацией вещества, и молекулы последнего могут связываться со специфическими участками. В результате конформационных изменений в белке участки связывания вместе с переносимым веществом экспонируются в раствор с низкой его концентрацией (состояние «пинг»). Этот процесс полностью обратим, и суммарный поток вещества через мембрану определяется его концентрационным градиентом. Скорость, с которой растворенное вещество поступает в клетку, зависит о г следующих факторов: 1) трансмембранного концентрационного градиента; 2) количества переносчика (ключ к регуляции); 3) быстроты связывания вещества с переносчиком; 4) быстроты конформационных изменений нагруженного и ненагруженного переносчика.
Гормоны регулируют облегченную диффузию, изменяя число доступных переносчиков. Инсулин повышает интенсивность транспорта глюкозы в жировых и мышечных тканях, индуцируя поступление новых переносчиков из некого внутриклеточного пула (см. рис. 51.13). Он также повышает транспорт аминокислот в печень и другие ткани. Одним из множества скоординированных эффектов глюкокортикоидных гормонов является повышение транспорта аминокислот в печень, где они служат субстратом глюконеогенеза. Гормон роста усиливает транспорт аминокислот во все клетки, а эстрогены стимулируют этот процесс в матке. В животных клетках существуют по меньшей мере пять разных систем переносчиков аминокислот. Каждая из них специфична к определенной группе близкородственных аминокислот и может функционировать как система симпорта с Na+ (рис. 42.13).
Б. Активный транспорт. Процесс активного транспорта отличается от диффузии тем, что он сопровождается смещением состояния системы от термодинамического равновесия и, следовательно, требует энергетических затрат. Источником энергии могут быть гидролиз АТР, процесс переноса электронов или свет. Поддержание электрохимических градиентов играет столь большую роль в биологических системах, что на него затрачивается около 30—40% всей потребляемой клеткой энергии.
В основном в клетках поддерживается низкая внутриклеточная концентрация Na+ и высокая К+ (табл. 42.1) и вместе с тем — суммарный отрицательный электрический потенциал. Насосом, который поддерживает эти градиенты, является АТРаза, активируемая ионами Na+ и К+ (рис. 42.17). Эта АТРаза — интегральный белок, для своей активности она требует фосфолипидов. Каталитические центры АТРазы для АТР и Na+ расположены на цитоплазматической стороне мембраны, а центр связывания К+ — на наружной. Уабаин ингибирует активность АТРазы, связываясь с ее внеклеточным фрагментом. Это ингибирование может частично сниматься внеклеточным К+.
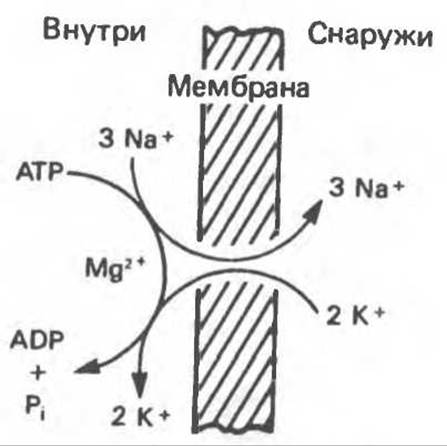
Рис. 42.17. Стехиометрия Na+, К+-насоса. Насос переносит три иона Na+ из клетки и два иона К+ в клетку на каждую молекулу АТР, гидролизуемую до ADP мембраносвязанной АТРазой. Уабаин и другие сердечные гликозиды блокируют насос при введении их во внеклеточную среду. (С любезного разрешения R. Post.)
Распространение нервного импульса
На мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраны электрически возбудимы. При химической стимуляции, опосредуемой специфическим синаптическим мембранным рецептором (см. разд. «Передача биохимических сигналов»), происходит срабатывание воротных механизмов, и в клетку быстро начинают поступать Na+ и Са2+ (при этом К+ может и не выходить из клетки), напряжение на мембране резко падает, и соответствующий ее участок оказывается деполяризованным, но в результате работы ионных насосов электрохимический градиент быстро восстанавливается.
Когда таким образом деполяризуются большие участки мембраны, электрохимическое возмущение распространяется вдоль мембраны подобно волне, порождая нервный импульс. Миелиновые оболочки, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят в клетку и выходят из нее только в тех местах, где изолятор отсутствует. Миелиновая мембрана состоит из фосфолипидов, в частности из сфингомиелина, холестерола, а также белков и гликосфинголипидов. С ней ассоциированы лишь немногие интегральные и периферические белки, которые, по-видимому, удерживают вместе многочисленные мембранные бислои, образующие гидрофобную изолирующую структуру, непроницаемую для ионов и воды. Некоторые заболевания, например рассеянный склероз и синдром Гиллайна-Барре, характеризуются демиелинизацией и нарушением проведения нервного импульса.
Транспорт глюкозы
На примере транспорта глюкозы мы сможем суммировать ряд ключевых положений, приведенных в этой главе. Транспорт глюкозы в клетку — это первый этап утилизации энергии. Исключением из общего правила является печень, в которой такой специфический процесс обнаружен не был. В клетки печени глюкоза поступает путем простой диффузии по концентрационному градиенту, который всегда чрезвычайно велик из-за быстрого превращения внутри клетки глюкозы в глюкозо-6-фосфат. В другие клетки (жировые и в еще большей степени мышечные) глюкоза поступает с помощью специфической транспортной системы, регуляция которой осуществляется инсулином (см. рис. 51.13). Изменения транспорта обусловлены в основном изменением Vmax (преимущественно благодаря увеличению или уменьшению числа переносчиков), но могут быть связаны и с вариациями Kм. Структура переносчика глюкозы эритроцитов была определена по последовательности соответствующей кДНК (рис. 42.7). Трансформированные с помощью к ДНК клетки синтезируют этот белок и встраивают его в мембрану в функционально активном состоянии; дальнейшие исследования с помощью направленного мутагенеза, возможно, помогут выяснить, как функционирует этот белок.
Рассматривая транспорт глюкозы, мы сталкиваемся с различными аспектами транспорта веществ, рассмотренными выше. Глюкоза и Na+ связываются с разными участками переносчика глюкозы. При этом Na+ поступает в клетку под действием электрохимического градиента и «тащит» глюкозу за собой (рис. 42.18). Таким образом, чем круче градиент Na+, тем больше поступает глюкозы, и, если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы подавляется. Чтобы поддерживать необходимый для работы переносчика Na+/глюкозы градиент Na+, используется Na+, K+-насос, поддерживающий низкую внутриклеточную концентрацию Na+. Аналогичные механизмы используются клетками для транспорта других сахаров, а также аминокислот.
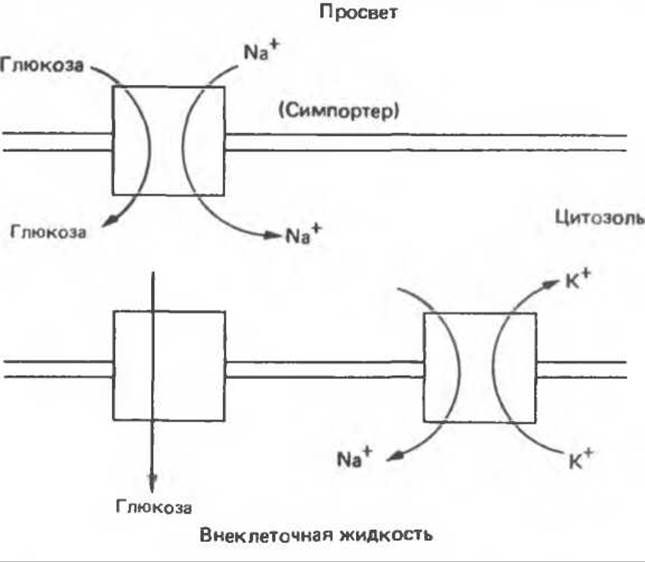
Рис. 42.18. Трансклеточное перемещение глюкозы через клетку кишечника. Через эпителиальную мембрану с люминальной стороны глюкоза проходит вслед за Na+. Градиент Na+. являющийся движущей силой этого симпорта, создается в процессе Na+, К+-обмена через базальную мембрану, обращенную к внеклеточной жидкости Глюкоза, сконцентрированная в клетке, перемещается затем по градиенту во внеклеточную жидкость с помощью облегченной диффузии (по механизму унипорта)
Трансклеточное перемещение сахаров включает один дополнительный компонент — унипортер, с помощью которого глюкоза, поступившая в клетку через одну ее поверхность, может выходить через другую; такой процесс наблюдается в клетках почек и кишечника.