Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Мембраны: структура, сборка и функции
Трансмембранное перемещение макромолекул
Через плазматическую мембрану транспортируются также макромолекулы. Процесс, с помощью которого клетки захватывают крупные молекулы, называется эндоцитозом. Некоторые из этих молекул (например, полисахариды, белки и полинуклеотиды) служат источником питательных веществ. Эндоцитоз позволяет также регулировать содержание определенных мембранных компонентов, в частности рецепторов гормонов. Эндоцитоз можно использовать для более детального изучения клеточных функций. Клетки одного типа можно трансформировать с помощью ДНК другого типа и, таким образом, изменить характер их функционирования или фенотип. В таких экспериментах часто используют специфические гены, что предоставляет уникальную возможность изучать механизмы их регуляции. Трансформация клеток с помощью ДНК осуществляется путем эндоцитоза — именно таким способом ДНК поступает в клетку. Трансформацию обычно проводят в присутствии фосфата кальция, поскольку Са2+ стимулирует эндоцитоз и осаждение ДНК, что облегчает ее проникновение в клетку с помощью эндоцитоза. Из клетки макромолекулы выходят путем экзоцитоза. Как при эндоцитозе, так и при экзоцитозе образуются везикулы, сливающиеся с плазматической мембраной или отшнуровывающиеся от нее.
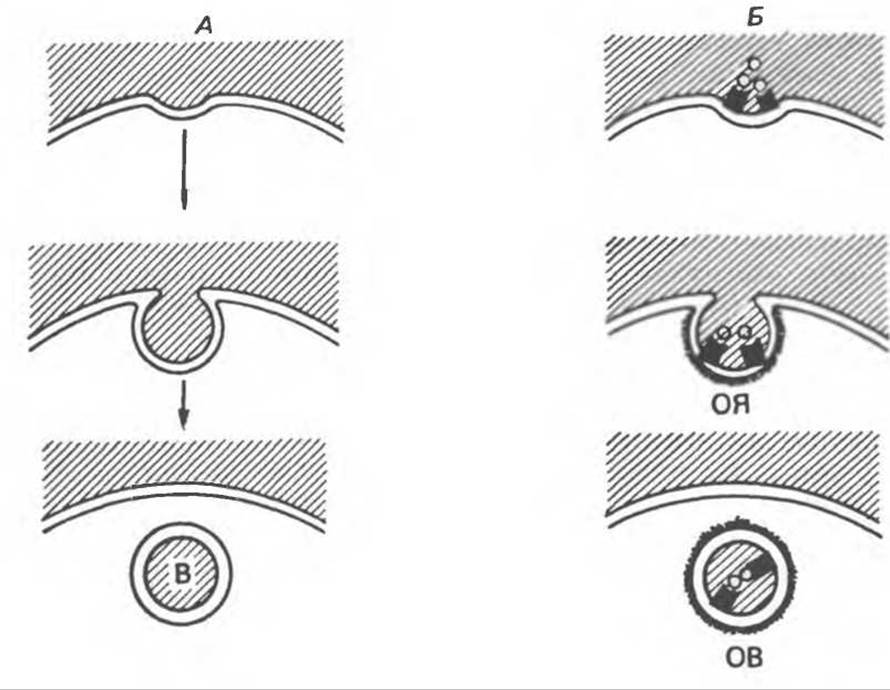
Рис. 42.19. Два типа эндоцитоза. Эндоцитозные везикулы (В) образуются в месте инвагинации плазматической мембраны. Жидкофазный пиноцитоз (А) — это случайный процесс, не имеющий определенной направленности. Опосредованный рецептором пиноцитоз (Б) селективен и осуществляется путем образования окаймленных ямок (ОЯ), выстланных белком клатрином (аморфное вещество), и окаймленных везикул (OB). Его специфичность обеспечивается рецепторами (черные прямоугольники), специфичными для разных молекул.
Эндоцитоз
У всех эукариотических клеток часть плазматической мембраны постоянно оказывается внутри цитоплазмы. Это происходит в результате инвагинации фрагмента плазматической мембраны, образования эндоцитозной везикулы, замыкания шейки везикулы и отшнуровывания ее в цитоплазму вместе с содержимым (рис. 42.19). Впоследствии везикулы могут сливаться с другими мембранными структурами и, таким образом, переносить свое содержимое в другие клеточные компартменты или даже обратно, во внеклеточное пространство. Большинство эндоцитозных везикул сливаются с первичными лизосомами и образуют вторичные лизосомы, которые содержат гидролитические ферменты и являются специализированными органеллами. Макромолекулы перевариваются в них до аминокислот, простых сахаров и нуклеотидов, которые диффундируют из везикул и утилизуются в цитоплазме. Для эндоцитоза необходимы: 1) энергия, источником которой обычно служит АТР; 2) внеклеточный Са2+; 3) сократительные элементы в клетке (вероятно, системы микрофиламентов).
Эндоцитоз можно подразделить на два основных типа. Фагоцитоз осуществляется только с участием специализированных клеток, таких, как макрофаги и гранулоциты. При фагоцитозе происходит поглощение крупных частиц — вирусов, бактерий, клеток или их обломков. Макрофаги исключительно активны в этом отношении и могут включать в себя объем, составляющий 25% собственного объема, за 1 ч. При этом происходит интернализация 3% их плазматической мембраны каждую минуту, или целой мембраны каждые 30 минут.
Пиноцитоз присущ всем клеткам. С его помощью клетка поглощает жидкости и растворенные в ней компоненты. Этот процесс также можно подразделить на два типа. Жидкофазный пиноцитоз — это неизбирательный процесс, при котором количество растворенного вещества, поглощаемого в составе везикул, просто пропорционально его концентрации во внеклеточной жидкости. Такие везикулы образуются исключительно активно. Например, у фибробластов скорость интернализации плазматической мембраны составляет 1 /3 скорости, характерной для макрофагов. В этом случае мембрана расходуется быстрее, чем синтезируется. В то же время площадь поверхности и объем клетки сильно не меняются, что указывает на восстановление мембраны за счет экзоцитоза или за счет повторного ее включения с той же скоростью, с какой она расходуется.
Другой тип пиноцитоза, адсорбционный пиноцитоз, представляет собой селективный процесс, опосредуемый медиатором. Он ответствен в основном за поглощение макромолекул, для которых на плазматической мембране существует ограниченное число связывающих участков. Эти рецепторы, обладающие высоким сродством, выборочно концентрируют лиганды из среды при минимуме поглощаемой жидкости и растворенных в ней несвязывающихся молекул и заметно увеличивают эффективность поступления специфических молекул в клетку. Везикулы, образующиеся при адсорбционном пиноцитозе, образуются в месте инвагинаций (ямок), покрытых с цитоплазматической стороны волокнистым материалом. Обычно таким материалом является клатрин (вероятно, периферический мембранный белок). Окаймленные ямки могут занимать до 2% поверхности некоторых клеток.
С помощью окаймленных ямок, в которых располагаются соответствующие рецепторы, интернализуются, например, липопротеины низкой плотности (ЛНП) и их рецепторы (см. гл. 26). Эндоцитозные везикулы, содержащие ЛНП и их рецепторы, сливаются в клетке с лизосомами. Рецепторы освобождаются и возвращаются на поверхность клеточной мембраны, а апопротеин ЛНП расщепляется и соответствующий эфир холестерола метаболизируется. Синтез рецепторов ЛНП регулируется вторичными или третичными продуктами пиноцитоза, т.е. веществами, образующимися при метаболизме ЛНП, например холестеролом. Нарушения процессов образования рецептора ЛНП и его интернализации имеют большое биомедицинское значение (гл. 26).
С помощью адсорбционного пиноцитоза происходит поглощение и других макромолекул, в том числе некоторых гормонов. При этом образуются рецептосомы — везикулы, которые не сливаются с лизосомами и освобождают свое содержимое в другие внутриклеточные компартменты, например аппарат Гольджи.
Для осуществления адсорбционного пиноцитоза внеклеточных гликопротеинов необходимо, чтобы последние содержали специфический углеводный остаток, подлежащий распознаванию. Такие сигнальные остатки связываются с молекулами мембранного рецептора, который выполняет ту же функцию, что и рецептор ЛНП. На поверхности гепатоцитов находится галактозильный рецептор, с помощью которого осуществляется адсорбционный пиноцитоз сиалогликопротеинов. Кислая гидролаза, поглощаемая фибробластами посредством адсорбционного пиноцитоза, распознается благодаря маннозо-6-фосфатному остатку. Интересно, что этот остаток, по-видимому, играет важную роль в целевом перемещении гидролаз внутри клетки к лизосомам (см. гл. 54).
Опосредованный рецепторами эндоцитоз имеет свою теневую сторону, поскольку вирусы, вызывающие некоторые заболевания, например гепатит (повреждение клеток печени), полиомиелит (повреждение моторных нейронов) и СПИД (повреждение Т-клеток), атакуют клетки именно по этому механизму. Токсический эффект железа тоже начинает проявляться в результате его избыточного поглощения, обусловленного эндоцитозом.
Экзоцитоз
Большинство клеток высвобождают макромолекулы во внешнюю среду путем экзоцитоза. Этот процесс играет роль и в обновлении мембраны, когда ее компоненты, синтезированные в аппарате Гольджи, доставляются в составе везикул к плазматической мембране. Сигнал к началу эндоцитоза часто подается с помощью гормона, который, связываясь с рецептором на клеточной поверхности, индуцирует локальные и обратимые изменения концентрации Са2+, которые инициируют эндоцитоз. На рис. 4.20 схематически представлены процессы экзо- и эндоцитоза.

Рис. 42.20. Сравнение механизмов эндоцитоза и экзоцитоза. При экзоцитозе происходит слияние двух внутренних находящихся со стороны цитоплазмы монослоев, тогда как при эндоцитозе сливаются внешние монослои.
Вещества, высвобождаемые путем экзоцитоза, можно разделить на три категории: 1) вещества, связывающиеся с клеточной поверхностью и становящиеся периферическими белками, например антигены; 2) вещества, включающиеся во внеклеточный матрикс, например коллаген и глюкозаминогликаны; 3) вещества, выходящие во внеклеточную среду и служащие сигнальными молекулами для других клеток. Инсулин, паратиреоидный гормон и катехоламины упаковываются в гранулы и созревают внутри клетки, а затем при соответствующей стимуляции высвобождаются наружу (гл. 47, 49 и 51).