Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Гормоны гипофиза и гипоталамуса
Гормоны гипоталамуса
Секреция (и в некоторых случаях образование) каждого из гипофизарных гормонов, перечисленных в табл. 45.1, находится под тоническим контролем по меньшей мере одного гормона гипоталамуса. Гормоны гипоталамуса высвобождаются из окончаний гипоталамических нервных волокон, окружающих капилляры гипоталамо-гипофизарной системы в ножке гипофиза, и достигают передней его доли через специальную портальную систему сосудов, соединяющую гипоталамус и эту долю. Сведения о структуре некоторых гормонов гипоталамуса можно почерпнуть из табл. 45.2.
Таблица 45.1. Гипоталамо-гипофизарные гормоны
|
Гормоны гипоталамуса |
Сокращение |
Высвобождаемый гормон гипофиза1) |
|
Кортикотропин-рилизинг-гормон (кортиколиберин) |
КРГ |
АКТГ (ЛПГ, МСГ, эндорфины) |
|
Тиреотропин-рилизинг-гормон (тиролиберин) |
ТРГ |
ТТГ |
|
Гонадотропин-рилизинг-гормон (гонадолиберин) |
ГнРГ |
Л. ФСГ |
|
Гормон роста-рилизинг-гормон (соматолиберин) |
СТГ-РГ |
ГР |
|
Гормон, ингибирующий высвобождение гормона роста (соматостатин) |
СС |
ГР (ТТГ, ФСГ, АКТГ) |
|
Гормоны, ингибирующие высвобождение пролактина; дофамин и ГАП |
ПИГ (пролактин-ингибирующий гормон) |
Пролактин |
1) В скобках—гормоны гипофиза, на высвобождение которых данный гипоталамический гормон оказывает вторичное или более слабое действие.
Гипоталамические гормоны высвобождаются в пульсирующем режиме, и изолированные клетки- мишени передней доли гипофиза лучше реагируют на пульсовое введение этих гормонов, чем на их длительное воздействие. Высвобождение лютропина (ЛГ) и фоллитропина (ФСГ) контролируется концентрацией одного и того же рилизинг-гормона, гонадолиберина, а его концентрация в свою очередь определяется уровнем в крови половых гормонов, достигающих гипоталамуса (см. петлю обратной связи на рис. 43.1). Высвобождение адренокортикотропина (АКТГ) контролируется в основном кортиколиберином (кортикотропин-рилизинг-гормоном, КРГ), но в регуляцию этого процесса может быть вовлечен и ряд других гормонов, включая антидиуретический гормон (АДГ), катехоламины, вазоактивный интестинальный пептид (ВИП) и ангиотензин II. На секрецию кортиколиберина влияет кортизол (глюкокортикоидный гормон, секретируемый надпочечниками). Высвобождение тиреотропина (ТТГ) зависит главным образом от тиролиберина (тиреотропин-рилизинг-гормон, ТРГ), секреция которого в свою очередь регулируется гормонами щитовидной железы, трииодтиронином (Т3) и тироксином (Т4); секреция ТТГ тормозится соматостатином (см. рис. 46.4). Секреция и продукция гормона роста (ГР) находятся под тоническим контролем как стимулирующих, так и ингибирующих гипоталамических гормонов. Кроме того, в регуляции секреции гормона роста участвует периферическая петля обратной связи. Инсулиноподобный фактор роста 1 (соматомедин С), который опосредует некоторые эффекты гормона роста, стимулирует высвобождение соматостатина и ингибирует секрецию соматолиберина (рис. 45.5). Регуляция синтеза и секреции пролактина (ПРЛ) сводится преимущественно к тоническому подавлению этих процессов гипоталамическими агентами. Ее отличительная особенность состоит в сочетании нервного (раздражение грудных сосков) и нейромедиаторного/нейрогормонального факторов. Дофамин (табл. 45.2) тормозит синтез пролактина (ингибируя транскрипцию пролактинового гена) и его секрецию; однако существуют данные, свидетельствующие о том, что подавление секреции пролактина обусловлено действием не одного дофамина. Недавно открыт 56-членный нейропептид, обладающий как гонадолибериновой активностью, так и активностью гормона, подавляющего высвобождение пролактина (пролактостатина). Его называют гонадолиберин-ассоциированным (связанным) пептидом (ГАП, GAP). ГАП, общая структура которого и ее специфические особенности представлены на рис. 45.1, является мощным ингибитором секреции пролактина, и его, очевидно, можно считать пролактостатином. Существованием ГАП можно объяснить любопытную связь между секрецией гонадолиберина и пролактина, которая особенно выражена у некоторых видов.
Таблица 45.2. Структура рилизинг-гормонов (либеринов) гипоталамуса
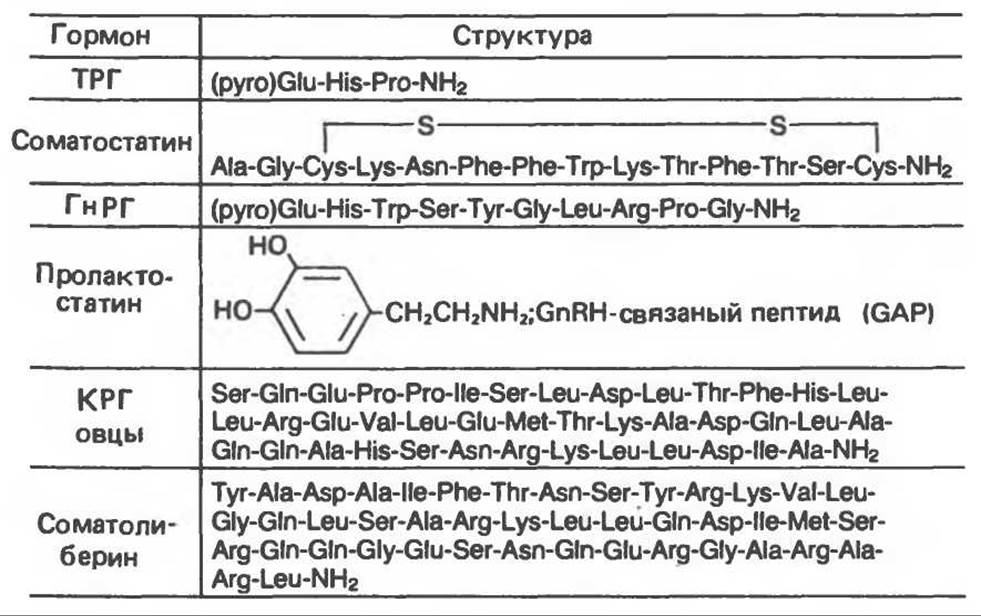

Рис. 45.1. Структура и аминокислотная последовательность плацентарной кДНК для препрогонадотропин-рилизинг-гормона (ГнРГ). Белок состоит из трех доменов: сигнального пептида, ГнРГ и гонадолиберин-ассоциированного пептида (ГАП). В нижней части рисунка представлены аминокислотные последовательности ГнРГ и ГАП. Указан сайт ферментативного процессинга, приводящего к разделению двух молекул. Цифры обозначают положения в аминокислотной последовательности ГнРГ (1—10) и ГАП (1—56). (Reproduced with permission, from Nicolics К. et. al. A prolactin-inhibiting factor with the precursor for human gonadotropin-releasing hormone. Nature 1986, 316, 511. Copyright 1985 by Macmillan Journals Ltd.)
Многие гипоталамические гормоны, в частности тиролиберин, кортиколиберин и сомагостатин, обнаруживаются в других отделах нервной системы и в ряде периферических тканей. Концентрация соматостатина в поджелудочной железе выше, чем в гипоталамусе. Он образуется D-клетками островков Лангерганса и, по-видимому, регулирует секрецию глюкагона и инсулина. Кроме того, соматостатин входит в число более чем 40 пептидов, продуцируемых нейронами центральной и периферической нервной системы.
Посредником действия рилизинг-гормонов на аденогипофиз первоначально считали сАМР, однако недавние опыты с гонадолиберином и тиролиберином позволяют предположить участие в соответствующих процессах кальций-фосфолипидного механизма, подобного описанному выше (см. рис. 44.5). Вопрос о том, влияют ли рилизинг-гормоны помимо секреции и на синтез соответствующих гормонов гипофиза, остается спорным; недавно было показано, что соматолиберин повышает скорость транскрипции гена гормона роста, а тиролиберин оказывает аналогичное действие на ген пролактина.