Биохимия - Химические реакции в живой клетке Том 1 - Д. Мецлер 1980
Молекулы, из которых мы состоим
Нуклеиновые кислоты
Двойные спирали
Одно из наиболее впечатляющих открытий в биологии в нашем веке было сделано в 1953 г., когда Джеймс Уотсон и Фрэнсис Крик установили, что ДНК представляет собой двойную спираль, составленную из двух антипар аллельных полинуклеотидных цепей1. Наиболее важной особенностью предложенной структуры было спаривание оснований противоположных цепочек путем образования между ними водородных связей. Водородные связи (на рис. 2-21 они указаны пунктирными стрелками) могут образоваться лишь в том случае, если всюду вдоль структуры ДНК аденин образует пару с тимином (две водородные связи), а цитозин — с гуанином (три связи). Таким образом, последовательность нуклеотидов в одной цепи оказывается комплементарной, но не идентичной последовательности в другой цепи. Далее почти сразу же стало очевидно, что последовательность оснований в цепи ДНК содержит в себе закодированную генетическую информацию. Комплементарность двух цепей приводит к очень простому механизму репликации генов на протяжении всех клеточных делений. По этому механизму две цепи ДНК разделяются и вдоль каждой из них синтезируется новая комплементарная цепь, что дает в результате две молекулы ДНК, по одной на каждую из двух дочерних клеток. Принципиальную правильность этой схемы сейчас уже можно считать доказанной.

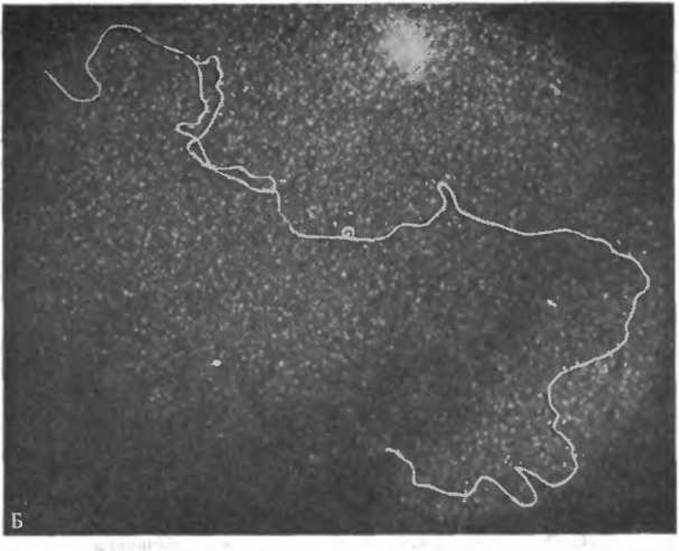
РИС. 2-23. А. Двойная спираль ДНК; В-форма. (Arnott S., Hukins D. W. L., JMB, 81, 93—105, 1975.) Б. Электронная микрофотография молекулы ДНК бактериального вируса (бактериофаг Т7) в момент ее репликации. Вирусная ДНК представляет собой длинный (~14 мкм) дуплексный стержень, содержащий около 40 000 пар оснований. Виден небольшой репликативный «глаз» — участок, где происходит удвоение ДНК. Синтез ДНК начинается в особой точке (точке инициации), расположенной иа расстоянии, равном 17% длины молекулы, от одного из концов дуплекса. Окраска уранилацетатом; негативное контрастирование. (С любезного разрешения Т. Wolfson и D. Dressier.)
Геометрия двойной спирали представлена на рис. 2-23,21. В волокнах молекулы ДНК могут находиться в двух конформациях: А и В. A-форма1 характеризуется тем, что плоскости пар оснований отклонены от нормали к оси спирали примерно на 20°, тогда как в В-форме, реализуемой при высокой влажности, плоскости почти перпендикулярны оси спирали (торсионные углы остова составляют ω = 155°, ξ = 36°, θ = —146°, φ = —96° и ψ = 46°) [65] Считается, что ДНК внутри клеток находится в основном в В-форме.
1 Эта структура была найдена в результате построения молекулярных моделей с привлечением рентгеновских дифракционных данных, полученных Уилкинсом и Франклин при исследовании ориентированных волокон ДНК. За это открытие Уотсон, Крик н Уилкинс в 1962 г. были удостоены Нобелевской премии.
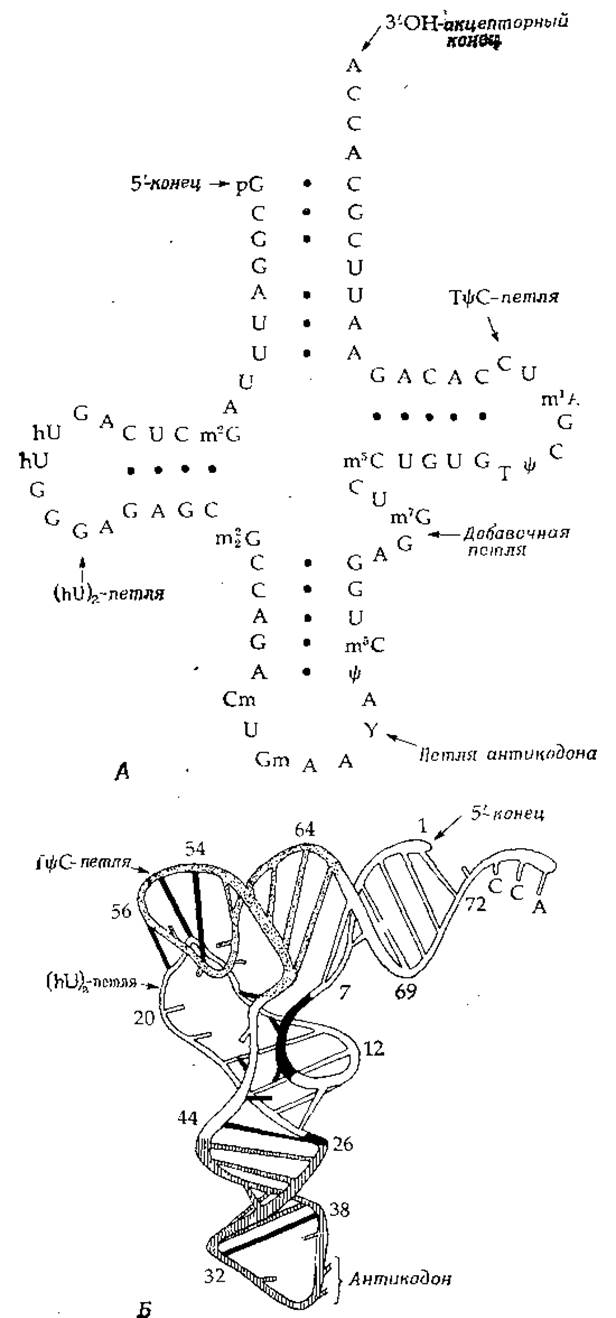
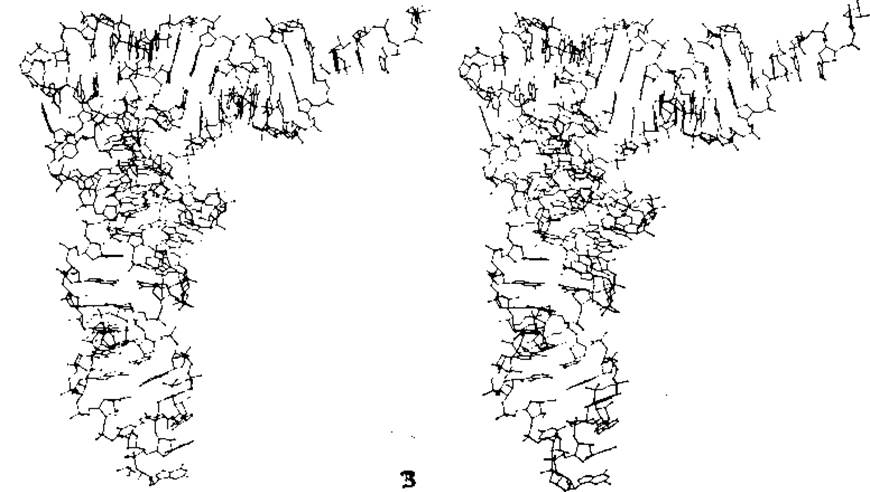
РИС. 2-24. Структура фенилаланиновой транспортной РНК из дрожжей. А. Последовательность нуклеотидов, образующая традиционную структуру «клеверного листа». Б. Схема пространственной укладки полинуклеотидной цепи. Рибозофосфатный остов изображен в виде протяженной непрерывной ленты; прямолинейные полосы, соединяющие разные участки ленты, — это пары оснований, связанные водородными связями. Одиночные основания изображены в виде укороченных полосок. ТψС-петля выделена точками, литикодоновая петля — вертикальной штриховкой. Взаимодействия на уровне третичной структуры отмечены черными полосками. (Quigley G. J., Rich A., Science, 194, 796—806, 1976.) В. Стереоизображение структуры дрожжевой фенилаланиновой тРНК по данным рентгеноструктурного анализа. Акцепторный участок с выступающей 3'-коицевой последовательностью АССА расположен справа, антикодон GAA — в правой нижней части рисунка. Особенно хорошо видно гуаниновое кольцо в самой нижней части рисунка. Плоскость среднего аденина антикодона расположена строго перпендикулярно плоскости рисунка, как и для «гипермодифицированного» основания Y (см. рис. 15-10), которое расположено непосредственно над антикодоном. Его боковая цепь находится в дальней от нас части изображения. Непосредственно перед антикодоном, в 5'-«половине» молекулы, расположены два неспаренных основания (С и U). 2'-гидроксильные группы цитидина и входящего в состав антикодона гуанозина метилированы. Двигаясь вверх вдоль петли антикодона, мы проходим два триплета оснований, в которых реализуется схема спаривания как Уотсона — Крика, так и Хугстена. Рекомендации относительно того, как рассматривать такие рисунки, даны в подписи к рис. 2-6. (С любезного разрешения A. Rich.)
Если пренебречь асимметрией пары оснований, то можно заметить, что в составе двойной спирали одна нуклеотидная цепь связана с другой осью симметрии 2-го порядка. Этот элемент симметрии, порождаемый антипараллельным расположением цепей, делает молекулу ДНК с обоих концов одинаковой — как с точки зрения человека, рассматривающего модель, так и с точки зрения фермента, вступающего с молекулой во взаимодействие. В действительности две цепи не идентичны, а генетическая информация может считываться с тех участков, которые располагаются на поверхности в районе большой бороздки (рис. 2-23, А).
1 В действительности мы имеем здесь дело с семейством близких форм.
Приведем некоторые данные о размерах двойной спирали (рис. 2-23). Диаметр ее, определяемый расстоянием между атомами фосфора, равен в точности 2,0 нм. Шаг спирали — 3,4 нм, на один виток приходится десять пар оснований. Таким образом, расстояние между плоскостями оснований равно 0,34 нм, что примерно эквивалентно сумме вандерваальсовых радиусов ароматических колец (табл. 2-1). Таким образом, основания уложены в виде стопки в центре спирали. Типичный ген в 1000 оснований — это участок ДНК около 340 нм длиной (рис. 2-23, Б).
Как и подобает матрице, с которой снимаются копии, двойная спираль ДНК исключительно стабильна. Несмотря на большую длину, в природных условиях она расщепляется крайне редко. Такая стабильность структуры обусловлена несколькими факторами: 1) наличием водородных связей между основаниями; 2) вандерваальсовым притяжением плоских оснований, уложенных параллельно одно над другим; 3) присутствием на поверхности молекулы многих атомов кислорода, отрицательно заряженных и нейтральных, способных к образованию водородных связей с водой или со специфическими белками, окружающими молекулу; 4) способностью к образованию различного рода суперспиралей (см. ниже).
Как мы отмечали выше, ДНК может переходить в паракристаллическую A-форму (с наклонным расположением оснований и 11 парами оснований на виток). Напрашивается вывод, что и в природных условиях эта конформация не менее важна, чем В-форма. Хотя молекулы РНК обычно одноцепочные, они часто образуют шпильки — двухцепочечные участки, находящиеся в А-форме [71]. В-конформация в этом случае исключается присутствием 2'-гидроксильных групп в рибозе РНК. Считается, что в клетках образуется также и переходная «гибридная» двойная спираль, составленная из молекул ДНК. и РНК, которая, по всей видимости, тоже ограничена рамками A-формы. Следует отметить, что A-форма отличается от В-формы еще и тем, что имеет довольно большую (~0,8 нм в диаметре) полость вдоль оси спирали, а большая бороздка у нее более глубокая [71а]. В отличие от структуры, изображенной на рис. 2-23, в A-форме плоскости пар оснований не пересекают оси спирали.
Наиболее тщательно изучена структура низкомолекулярной тРНК. Во всех этих молекулах существуют двухцепочечные участки, стабилизированные водородными связями с образованием трех шпилек, к которым иногда добавляется четвертая («клеверный лист»). Структура одной из тРНК установлена методом рентгеноструктурного анализа [72—74] (рис. 2-24). Нерегулярность и сложность формы молекулы ставит ее в один ряд с молекулами глобулярных белков. Обратите внимание на расположенный в нижней части рисунка антикодон (триплет оснований), структура которого обеспечивает спаривание с тремя основаниями кодона, детерминирующего определенную аминокислоту, в данном случае фенилаланин.