Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Коферменты - особые природные специализированные реагенты
Коферментные формы витамина В12
Рибонуклеотидредуктаза
Рибонуклеотиды восстанавливаются до дезоксирибонуклеотидов, необходимых для синтеза ДНК, особыми ферментами, действующими на ди- и трифосфаты пуриновых и пиримидиновых нуклеозидов (гл. 14, разд. Л,1,а). У некоторых организмов для восстановления необходим витамин В12. Так, например, у Lactobacillus leichmanii трифосфаты восстанавливаются путем переноса водорода на дитиолы, например на дигидролипоевую кислоту или на дитиоловый белок, тиоредоксин (табл. 8-6). Протоны воды обратимо включаются в 2'-положение восстановленного нуклеотида с сохранением конфигурации. Эти данные указывают на то, что С-2'-атом водорода может переноситься через 5'-дезоксикобаламин в —SH-группу тиоредоксина, обменивающуюся со средой [174, 175].
Реакция редуктазы с дигидролипоевой кислотой в присутствии dGTP (который, по-видимому, выступает в роли аллостерического активатора) приводит к быстрому (на протяжении нескольких миллисекунд) образованию радикала с характерным ЭПР-спектром, который можно изучить, если реакционную смесь быстро охладить до 130 К. Если вместо dGTP использовать GTP (истинный субстрат), то сигнал радикала достигает максимального значения примерно через 20 мс, а затем уменьшается.
Из всех состояний окисления кобальта (3+, 2+ и 1+) только состояние 2+ витамина В12гявляется парамагнитным и имеет ЭПР-сигнал. Изменения электронного спектра поглощения кофермента рибонуклеотидредуктазы также подтверждают образование В12г. Таким образом, возможно гемолитическое расщепление В12г со стабилизацией 5'-дезоксиаденозильного радикала [175а]:
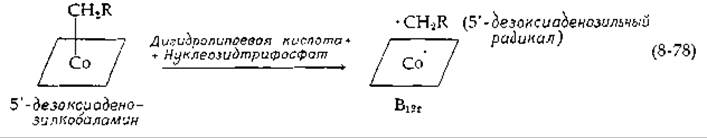
ЭПР-сигнал 5'-дезоксиаденозильного радикала не регистрируется. Поэтому не ясно, акцептируется ли атом водорода дигидролипоевой кислоты или нет. Также не ясно, каков механизм последующего стереоспецифического замещения 3'-ОН-группы рибонуклеотидного субстрата атомом водорода.
Вслед за рибонуклеотидредуктазой сходные сигналы ЭПР наблюдали и для других ферментов [176—178]. Таким образом, возможно, что во всех случаях начальной стадией является реакция (8-78). Если для реакций перегруппировки выполняется механизм (8-77), то стадию а следовало бы рассматривать как атаку субстрата со стороны либо восстановленного кобальта, либо 5'-дезоксиаденозильного радикала в уравнении (8-78).