Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Биосинтез; как образуются новые молекулы
Биосинтез мономеров
Биосинтез глюкозы (глюкогенез)
В предыдущих разделах мы рассмотрели пути биосинтеза трехуглеродных предшественников углеводов. Триозофосфаты образуются в восстановительном пентознофосфатном цикле (рис. 11-4,Б). Восстановительный цикл трикарбоновых кислот и глиоксилатный путь дают оксалоацетат, который легко превращается в фосфоенолпируват. Теперь мы рассмотрим дальнейшие превращения РЕР и триозофосфатов в глюкозо-1-фосфат — ключевое промежуточное соединение в биосинтезе всего обширного семейства сахаров и полисахаридов.
Процесс превращения РЕР в глюкозо-1-фосфат представляет собой обращение части гликолитической последовательности. Поэтому его имеет смысл рассматривать вместе с обращением всей последовательности гликолиза, начиная с молочной кислоты. Эта реакция, которая носит название глюконеогенеза, является важной составной частью цикла Кори (гл, 9, разд. Е). Она может быть использована организмом для превращения пирувата, образующегося в результате дезаминирования аланина или серина (гл. 14), в углеводы.
1) Интересно, что все гены, кодирующие специфические ферменты глиоксилатного пути, сосредоточены в одном участке бактериальной хромосомы.
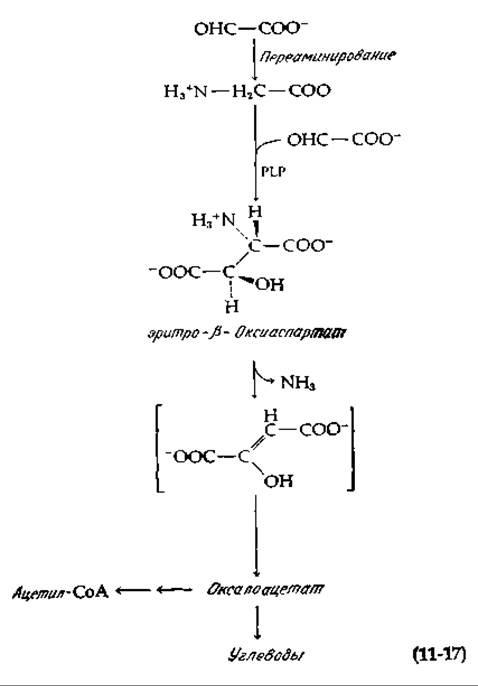
Так же, как и в случае пентозофосфатного цикла, точное обращение последовательности реакций гликолиза [уравнение (11-18)] термодинамически невыгодно.
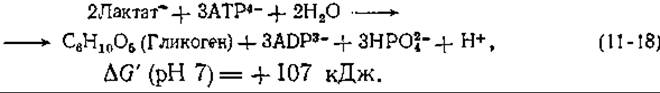
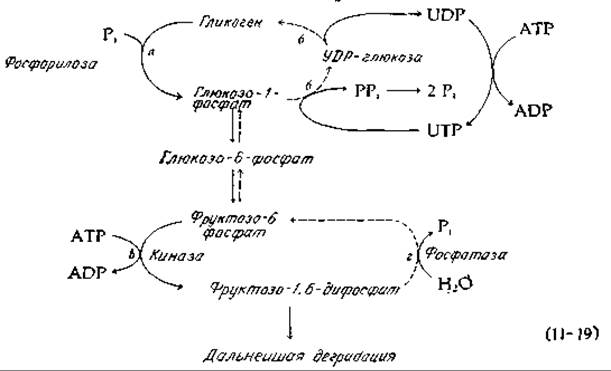
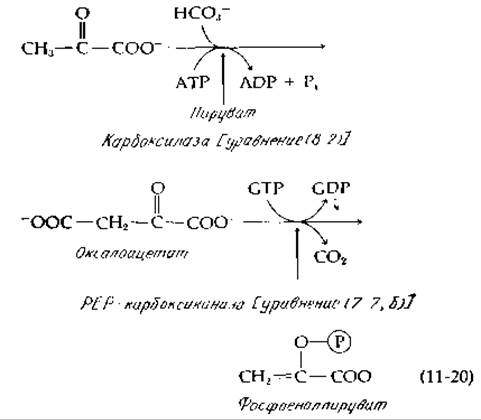
Даже при очень высоких значениях величины Rp маловероятно, чтобы реакция шла до конца. Действительные пути глюконеогенеза [уравнение 11-19), штриховые линии], по-видимому, отличаются от соответствующих путей в процессе гликолиза [уравнение (11-19), сплошные линии]. 1) Если расщепление гликогена инициируется реакцией неоргнического фосфата, катализируемой фосфорилазой [стадия а, уравнение (11-19)], то последовательность биосинтетических реакций от глюкозо-1-фосфата через глюкозоуридиндифосфат [уравнение (11-19), стадия б; см. также разд. Е,1,б] сопряжена с расщеплением молекулы АТР. 2) В катаболических процессах (гликолиз) фруктозо-6-фосфат превращается в фруктозо-1,6-дифосфат под действием специфической киназы [уравнение (11-19), стадия в], после чего образующийся фруктозодифосфат расщепляется альдолазой и образующийся триозофосфат претерпевает дальнейшее расщепление. В глюкогенезе же образование фруктозофосфата из фруктозодифосфата катализируется специфической фосфатазой [уравнение (11-19), стадия г]. 3) В процессе гликолиза фосфоенолпируват под действием киназы превращается в пируват с образованием АТР (рис. 9-7, реакция 10). В процессе же глюкогенеза пируват превращается в фосфоенолпируват не непосредственно, а через оксалоацетат [уравнение (11 -20)] А Это может служить другим примером сопряжения реакции расщепления АТР в последовательности карбоксилирование — декарбоксилирование. Суммарным результатом превращения пирувата в РЕР является использование не одной, а двух молекул АТР (фактически одной молекулы АТР и одной молекулы ГТР).
Суммарная реакция обращенного гликолиза характеризуется, таким образом, довольно большой отрицательной величиной изменения стандартной свободной энергии [уравнение (11-21)].
![]()
1) Однако общепринятое представление о том, что в процессе глюконеогенеза пируваткиназа не катализирует обратной реакции, было подвергнуто сомнению [17].
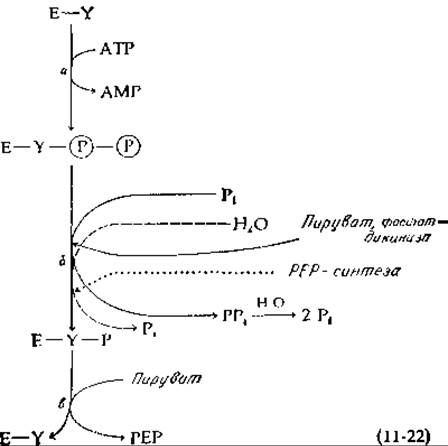
У бактерий и у растений были обнаружены два фермента, которые могут превращать пируват непосредственно в РЕР. В каждом из этих случаев, так же как и в рассмотренных в предыдущих разделах ферментных животных системах, такое превращение сопровождается расходованием двух высокоэнергетических связей АТР. Фосфоенолпируват- синтетаза Е. coli сначала, по-видимому, переносит пирофосфатную группу от АТР к Y-группе фермента [уравнение (11-22)], после чего одна фосфатная группа этого промежуточного соединения удаляется в результате гидролиза [пунктирная линия в уравнении (11-22), стадия б], обеспечивая образование большого количества промежуточного соединения E-Y-P, которое реагирует с пируватом и дает РЕР [18]. Аналогичный фермент, пируват, фосфат—дикиназа, был впервые идентифицирован в тропических травах, и сейчас известно, что он играет важную роль в функционировании систем, концентрирующих СО2 в так называемых С4-растениях (гл. 13, разд. Д,9) [19, 20]. Этот же фермент принимает участие в глюконеогенезе у Acetobacter [20]. О механизме функционирования этого фермента можно судить исходя из уравнения (11-22). Единственное его отличие от фосфоенолпируватсинтетазы Е. coli состоит в том, что атакующим нуклеофилом [уравнение (11-22), стадия в] является не вода, а неорганический фосфат, и в качестве продукта образуется неорганический пирофосфат, который, вероятно, подвергается затем гидролизу под действием фосфатазы. Суммарная реакция в конечном счете оказывается такой же, как и в случае РЕР-синтетазной реакции.
Здесь имеет смысл задать студенту вопрос. Дело в том, что такой же конечный результат должен был бы получиться, если бы промежуточное соединение Е—Y—Р в уравнении (11-22) образовалось в результате переноса фосфатной группы от АТР на Y. Продуктом явился бы в этом случае ADP. Для того чтобы довести процесс до конца, эту молекулу ADP можно было бы гидролизовать до АМР и Pi специальной фосфатазой. Почему этот процесс никогда не используется в природе?