Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Ферменты: белковые катализаторы клеток
Регуляция ферментативной активности
Усиление регуляторных сигналов
Имеющее регуляторное значение изменение активности фермента часто усиливается при помощи каскадного механизма: первый фермент воздействует на второй, второй — на третий и т. д. Этот механизм обеспечивает быстрое появление больших количеств активной формы последнего фермента цепи. Примером каскадного механизма может служить механизм свертывания крови [89], представленный схематически на рис. 6-16. Мы видим последовательность, состоящую из пяти ферментов и начинающуюся с фактора XII, в которой каждый фермент активирует следующий путем отщепления небольшой части пептидной цепи (ограниченный протеолиз). На конечном этапе тромбин воздействует на фибриноген и, отщепляя небольшой пептид, превращает его в фибрин — специализированный белок, который спонтанно свертывается. Какие факторы препятствуют выходу каскадного механизма из-под контроля? Почему при небольшом кровоподтеке весь протромбин в нашем организме не превращается в тромбин и не происходит свертывания всей крови? Здесь, несомненно, имеет место та же ситуация, что и в случае сАМР, который быстро удаляется из системы с помощью специфического фермента: существуют механизмы удаления активированного фермента из каскадной последовательности, представленной на рис. 6-16. Помимо этого имеется специальная ферментная система, растворяющая сгусток крови при заживлении раны [89].
Для одной из каскадных ферментных систем механизм, останавливающий развитие каскадного эффекта, известен. Гликогенфосфорилаза мышц активируется каскадной последовательностью ферментов, которая включается в результате контролируемого вегетативной нервной системой высвобождения адреналина (гл. 16, разд. Б, 3). Связывание адреналина мембраной клетки приводит к высвобождению сАМР, активирующего протеинкиназу. Киназа катализирует фосфорилирование другого фермента — киназы фосфорилазы. В этот момент мышцы готовы к быстрому расщеплению гликогена. Однако непосредственным сигналом к инициации этого процесса является высвобождение ионов Са2+ в цитоплазму в ответ на импульсы, поступающие от двигательных нейронов (гл. 4, разд. Е, 1). Киназа фосфорилазы активируется ионами кальция и в присутствии последних катализирует превращение неактивной фосфорилазы b в активную фосфорилазу а. Спонтанный возврат в состояние покоя происходит в результате действия фосфатаз, катализирующих отщепление фосфорильных групп, которые были ранее присоединены к белкам при помощи киназ. Важную роль в процессе играет также фосфодиэстераза, расщепляющая сАМР, и кальциевый насос, который снижает концентрацию активирующих ионов кальция до относительно низкого уровня.
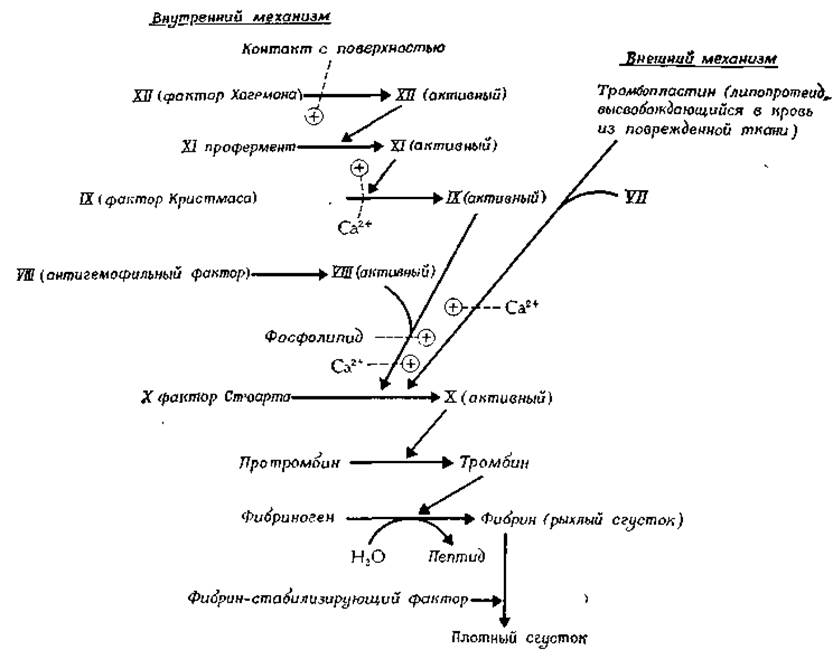
РИС. 6-16. Каскадный механизм, приводящий к свертыванию крови. Существуют два пути включения каскада: внутренний механизм запускается в результате контакта с поверхностью, внешний — в результате высвобождения тромбопластина из поврежденных тканей [89, 89а].
Еще один механизм усиления, детали которого только предстоит изучить, функционирует в сетчатке глаза. Известно, что одиночный квант света, падая на клетку рецептора, способен при подходящих условиях генерировать нервный импульс (гл. 16, разд. Б,3). Для распространения импульса необходимо, чтобы через мембрану прошло большое число ионов Na+, и вряд ли поглощение одного кванта света сможет инициировать фотохимическую реакцию, приводящую к интенсивному транспорту ионов натрия, если не произойдет соответствующего усиления сигнала.
Механизмы усиления могут иметь самую разную природу. Один из них, связанный с циклическим превращением субстрата, рассмотрен в гл. 11, разд. Е, 6.
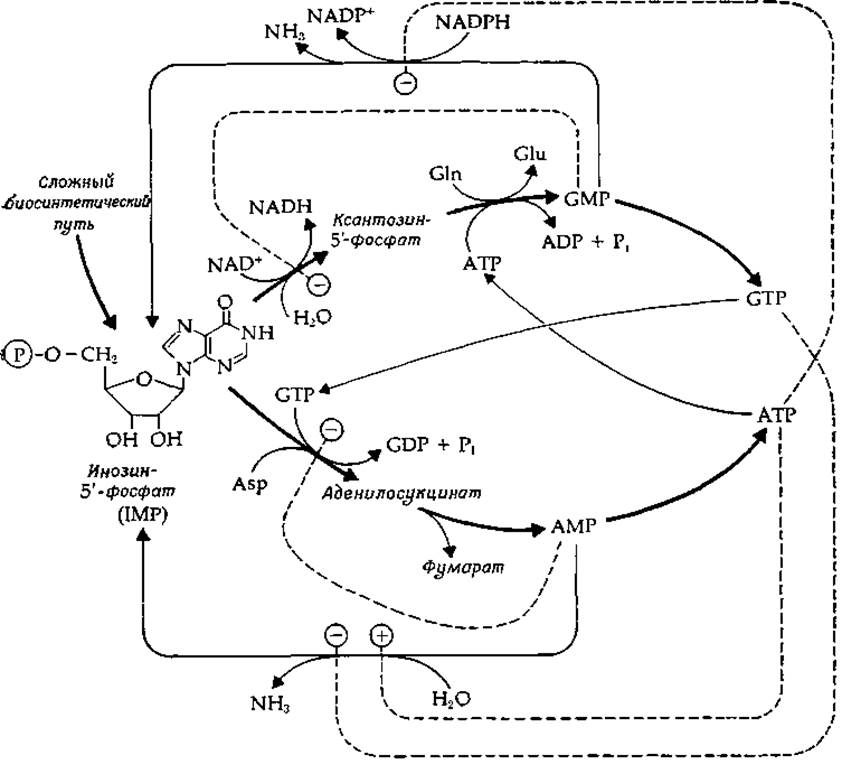
РИС. 6-17. Перекрестная регуляция синтеза пуриновых нуклеозидтрифосфатов (АТР и GTP) [66].