Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Хромосома эукариот и ее контроль
Организация
В геноме такого простого эукариота, как плесневый гриб Dictyostelium, содержится в 11 раз больше ДНК, чем в геноме Е. coli. У дрозофилы — «высшего» организма с наименьшим количеством ДНК—размер гаплоидного генома в 24 раза больше размера генома Е. coli. Кодирующая емкость генома человека в 600 раз больше, чем у бактерии (табл. 1-3). Столь большое количество ДНК является одной из причин, затрудняющих изучение эукариотического генома. Другая трудность обусловлена тем, что процесс транскрипции генов у эукариот может сильно изменяться как во времени, так и в зависимости от условий окружающей среды. Следовательно, механизмы регуляции фенотипического выражения генов должны быть очень сложными.
Геном высших организмов состоит из определенного числа отдельных хромосом, каждая из которых содержит, по-видимому, одну двухцепочечную молекулу ДНК. Эта молекула ДНК тесно связана с другими компонентами, в состав которых входит примерно 75% белка и 10% РНК (гл. 1, разд. Б,2). До недавнего времени мало что можна было сказать о том, как устроены хромосомы. Однако известно, что в профазе митоза или мейоза вытянутые хромосы иногда выглядят как нитки бус. Маленькие, богатые ДНК бусинки, известные под названием хромомер, подобно дискам политенных хромосом дрозофилы (разд. Г, 9, в), можно рассматривать как своего рода единицы генетической информации. Их существование дает основание думать, что ДНК в хромосоме каким-то образом разделена на отдельные единицы, возможно, аналогично оперонам бактерий.
Некоторые диски политенных хромосом выпячиваются и образуют пуфы. Самые крупные из них называют кольцами Бальбиани. В этих пуфах ДНК находится в наименее компактной форме; существует предположение, что именно здесь происходит активная транскрипция. Поскольку ДНК диска в политенной хромосоме содержит в среднем приблизительно 105 пар оснований, образуемые транскрипты мРНК могут быть очень большими [270]. В результате проведенных недавно исследований действительно было установлено, что одного кольца Бальбиани достаточно для образования 75S-PHK массой 15∙106—35∙106 дальтон [271]. Эта огромная молекула РНК переносится, по-видимому, в неизмененном виде целиком в цитоплазму, где происходит очень активный синтез белка. Возникает вопрос: является ли эта мРНК транскриптом одного гена и большого участка ДНК, не кодирующего синтез специфических белков, или же в ней заключена многократно повторяющаяся информация о синтезе какого-то одного белка? Этот вопрос имеет важное значение для понимания общих принципов - организации генетической информации в хромосомах эукариот. Не следует при этом забывать, что политенные хромосомы представляют особый тип модификаций, обнаруженный в клетках, подвергшихся окончательной дифференцировке и неспособных к размножению.
а. Хромосомы типа ламповых щеток
Другой особый вид хромосом, изучение которого позволило значительно расширить наши представления о ядрах эукариот, обнаруживается в профазе мейотического деления ооцитов. Речь идет о «хромосомах типа ламповых щеток», которые были подвергнуты детальному исследованию на амфибиях Xenopus. Хромосома типа ламповой щетки представляет собой гомологичную пару хромосом, каждая из которых в свою очередь состоит из двух тесно связанных друг с другом хроматид. Хромосомы находятся в предельно линейной, несконденсированной форме, причем около 5% содержащейся в них ДНК образует приблизительно 4000 точно спаренных петель, которые видны под электронным микроскопом. Каждая такая петля образована нитью двухцепочечной ДНК длиной около 50 нм, что соответствует приблизительно 150000 оснований. Тот факт, что ни в одной из петель не удается увидеть ни одного разрыва ДНК, подтверждает точку зрения, согласно которой единая молекула ДНК протянута в хромосоме через все петли от одного ее конца до другого.
Так же как и пуфы политенных хромосом (которые, возможно, имеют сходное строение), хромосомы типа ламповых щеток активно участвуют в транскрипции. Считают, что приблизительно 3% ДНК участвует в образовании мРНК, накапливающейся в ооците и функционирующей на ранних этапах эмбрионального развития [272]. Было бы логично предположить, что одна петля в хромосоме типа ламповых щеток, подобно одному диску политенной хромосомы, играет роль транскрипционной единицы. Однако здесь мы сталкиваемся со следующим парадоксом: количество ДНК, содержащееся в одном диске или в одной петле, достаточно для детерминирования 30—35 белков среднего размера. Тем не менее при анализе тонкой генетической структуры хромосомы дрозофилы в каждом диске удается обнаружить не более одной единицы комплементации [273]. Из этого следует, что всего лишь 3% ДНК дрозофилы содержат структурные гены для синтеза белков. Что же делает остальная ДНК и почему мутации в ней не приносят вреда организму? Ответы на эти вопросы до сих пор, к сожалению, не получены.
б. Повторяющиеся последовательности
Благодаря использованию химических методов детальная картина организации хромосом начинает вырисовываться более четко. В одном из типов экспериментов ДНК разрезают на фрагменты, включающие приблизительно по 10 000 пар оснований, и затем эти фрагменты денатурируют нагреванием. Оказалось, что при охлаждении ренатурация образовавшихся одноцепочечных фрагментов протекает по меньшей мере в два этапа. В одной части материала структура двойной спирали быстро восстанавливается, тогда как в другой части ренатурация протекает медленно (рис. 15-34) [273—275].
Как показывает опыт, быстро ренатурирующие фрагменты ДНК по нуклеотидному составу часто отличаются от основной массы ДНК. В результате при центрифугировании фрагментированной ДНК в градиенте плотности CsCl быстро ренатурирующая фракция имеет тенденцию отделяться, образуя узкую «сателлитную» область. Оказалось, что эта сателлитная ДНК состоит из часто повторяющихся коротких последовательностей [276, 277]. Например, ДНК сателлитной области кенгуровой крысы содержит повторяющуюся последовательность 5'-GGACACAGCG-3. На долю этой часто повторяющейся последовательности приходится 11% всей ДНК клетки. Сателлитная ДНК обычно находится в участках хромосомы, которые не расплетаются в телофазе подобно остальной ДНК. Функции сателлитной ДНК неизвестны. Было высказано предположение, что повторяющиеся последовательности появляются в процессе эволюции в результате неравного кроссинговера между сестринскими хроматидами во время митоза [232а]. При помощи рестриктирующих эндонуклеаз ДНК человека была разрезана на ряд фрагментов. Сравнение электрофоретических картин для фрагментов ДНК мужчин и женщин показало, что Y-хромосома содержит многократно (несколько тысяч раз) повторяющуюся в виде тандема нуклеотидную последовательность.
Согласно другим данным, некоторые из повторяющихся последовательностей распределены по всему геному случайно. Об этом свидетельствует, например, тот факт, что при ренатурации фрагментов ДНК дрозофилы образуются кольца ДНК, которые можно увидеть с помощью электронного микроскопа [278]. Кольца во-время ренатурации могут образовываться в результате фрагментации внутри повторяющихся последовательностей. В хромосомах Xenopus может содержаться около 25% таких повторяющихся последовательностей. Данные, полученные при электронной микроскопии, свидетельствуют о том, что случайная реассоциация фрагментов ДНК приводит к образованию двухцепочечных участков, содержащих повторяющиеся нуклеотидные последовательности, с одноцепочечными «хвостами». Последние обычно не спариваются, поскольку содержат уникальные последовательности, пришедшие из разных генов. У Xenopus повторяющиеся фрагменты ДНК включают приблизительно 300 нуклеотидов, а неповторяющиеся, или уникальные, фрагменты, расположенные между ними, — приблизительно 800 нуклеотидов [275].
Многое об организации ДНК можно узнать при исследовании РНК-транскриптов. При сравнении ядерной и цитоплазматической мРНК Dictyostelium были обнаружены значительные различия [279]. Цепь мРНК, когда она находится в ядре, включает приблизительно 1600 нуклеотидов, но в цитоплазме она укорачивается приблизительно до 1300 нуклеотидов. Как ядерная, так и цитоплазматическая мРНК содержат небольшие участки полиадениловой кислоты (разд. Б, 5). Путем ферментативного гидролиза можно выделить эти участки и определить их длину. Оказалось, что в ядерной мРНК эти участки включают приблизительно 25 остатков адениловой кислоты. Очевидно, эти участки поли (А) 25 транскрибируются с ДНК вместе с остальными участками мРНК, поскольку сама ДНК содержит около 15 000 последовательностей поли(dT)25. Этого числа участков поли(dТ) достаточно для того, чтобы каждый ген содержал хотя бы один такой участок. Сходные участки поли(dТ) были обнаружены в ДНК всех многоклеточных. Так, например, в ДНК дрозофилы оказалось около 6000 таких участков, что находится в соответствии с наличием у нее 5000 дисков политенных хромосом. В геноме млекопитающих содержится около 100 000 участков поли(dТ).
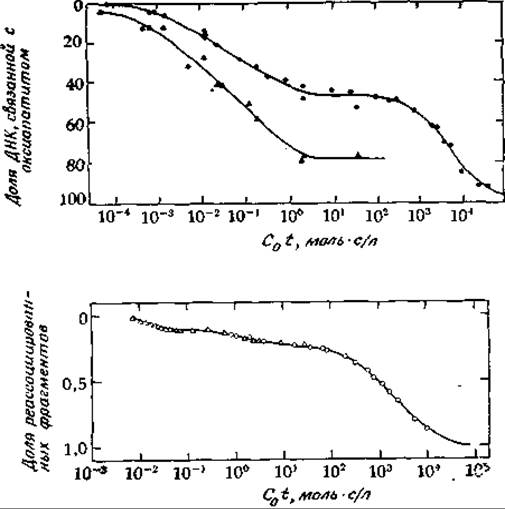
РИС. 15-34. Кривые ренатурации ДНК млекопитающих. А. Доказательство наличия в геноме теленка повторяющихся и неповторяющихся последовательностей. Верхняя кривая отражает реассоциацию фрагментов ДНК теленка, имеющих длину около 400 нуклеотидов. Инкубацию проводили при 60 °С в 0,12 М фосфатном буфере, а анализ проводился при тех же условиях оксиапатитным методом (гл. 2, разд. Г,10). Нижняя кривая отражает реассоциацию небольшого количества меченых фрагментов, содержащих 4000 нуклеотидов, с большим количеством фрагментов содержащих 400 нуклеотидов. ДНК последовательно фракционировали на три части: 1) быстро реассоциирующую фракцию (С0t≈3∙104), содержащую около 106 копий в геноме (сателлитная ДНК); 2) промежуточную фракцию (С0t≈0,01), содержащую около 66 000 копий (37%); 3) медленно реассоциирующую фракцию ДНК (С0t≈1000), не содержащую повторов (см. рис. 2-29 в статье Britten R. J., Smith J., Carnegie Inst. Wash. Yearb., 68, 378—391, 1970). Б. Реассоциация фрагментированной денатурированной ДНК мыши в 48%-ном растворе формамида и 5 х SSC (SSC означает раствор, содержащий 0,15 М NaCl и 0,015 М цитрата Na) при 37 °С. Реакцию реассоциации проводили при двух концентрациях: 1) 50 мкг/мл в стеклянных кюветах и с пришлифованными пробками с длиной оптического пути 1 см (∆) и 2) 1 мг/мл в кюветах с толщиной слоя 1 мм (О). Гипохромный эффект измеряли при 270 нм. Полученные данные приведены в виде зависимости количества ДНК, реассоциированной в течение данного времени, от произведения исходной концентрации ДНК (в молях фосфора на 1 л) на время реассоциации (в секундах). Установлено, что в образце содержится 10% быстро ренатурирующей сателлитной ДНК, 15% промежуточной частично измененной ДНК и 75% ДНК с уникальной последовательностью (McConaughy В. L., McCarthy В. J. Biochem. Genet, 4, 425—446, 1970).
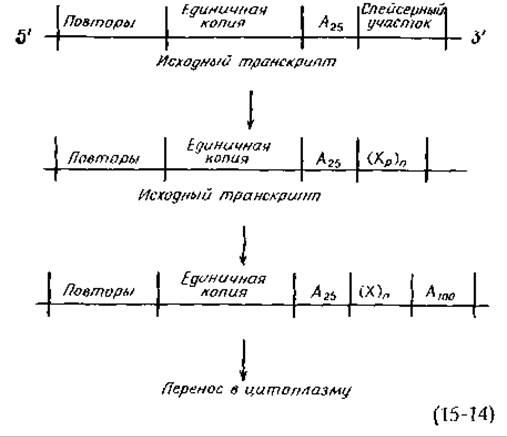
Что касается цитоплазматической мРНК, то, несмотря на меньший размер, она содержит больше поли (А). Так, например, в мРНК Dictyostelium кроме участков поли (А) 25 содержится такое же число более длинных участков поли (А), включающих приблизительно 100 остатков адениловой кислоты, которые добавляются, по-видимому, после транскрипции. Согласно рабочей гипотезе [уравнение (15-14)] [280], повторяющиеся последовательности, участки единичных копий, поли (А) 25 и «спейсерная область» имеются в каждом гене. После транскрипции от мРНК отщепляется кусок РНК, соответствующий спейсерному участку, оставляя на 3'-конце поли(А)25 неизвестное число нуклеотидов (Хр), и затем уже перед выходом в цитоплазму к ней присоединяется около 100 остатков адениловой кислоты [278]. Считают также, что могут иметь место другие «тандемные повторы», соответствующие «повторяющимся генам», которые для обеспечения клетки адекватными транскриптами должны существовать в виде больше чем одной копии.
в. Гены рибосомной РНК
Если большинство генов присутствует в хромосоме в единственном числе, то гены рибосомной РНК и тРНК представлены множеством копий. В случае Xenopus гены РНК 28S- и 18S-pибocoм повторяются в одной хромосоме приблизительно 450 раз. На концах длинных плеч большинства хромосом обнаруживается около 24000 копий генов- 5S-PHK [281]. Гены 28S- и 18S-PHK транскрибируются вместе с расположенным между ними спейсерным участком. Нетранскрибируемые спейсерные участки располагаются, как это видно на приведенных на рис. 15-11 электронных микрофотографиях, между повторяющимися парами генов. Известно, что для гена, кодирующего 5S-PHK, характерно высокое содержание GC-паp; следовательно, эти участки должны быть устойчивы к тепловой денатурации. Действительно, на денатурационной карте ДНК обнаруживаются легко денатурирующие участки, разделенные более короткими последовательностями из 120 оснований с высоким содержанием GC-пap, которые кодируют, вероятно, 5S-PHK. Легко денатурирующие (богатые АТ) спейсерные участки включают приблизительно 630 оснований. При помощи специфических рестриктирующих ферментов многие из этих АТ-богатых участков были разрезаны на повторяющиеся единицы, внутри которых также содержались повторы. Основной 15-нуклеотидный фрагмент с последовательностью. A4CUCA3CU3G повторялся примерно 30 раз [282].
В случае дрозофилы карта участка 5S-PHK в хромосоме 2, полученная при помощи рестриктирующих эндонуклеаз, показывает, что в этом участке содержатся два тандемно повторяющихся кластера генов и что в каждом из этих кластеров заключено около 90 генов. Как ориентированы кластеры, точно-не установлено, но существует предположение, что два кластера образуют длинный палиндром, переход которого в крестообразную конфигурацию, возможно, является одним из этапов в исправлении ошибок спаривания. Это способствовало бы сохранению гомогенности кластера [282а]. Данное соображение в равной мере применимо и в отношении большого числа длинных палиндромов, содержащихся в эукариотической ДНК [235а].
г. Амплификация генов
При определенных обстоятельствах часть генома может амплифицироваться путем повторной репликации одного или нескольких генов. Наиболее широко известным примером является амплификация генов рибосомной РНК ооцитов амфибий. В случае Xenopus избыток ДНК скапливается вокруг ядрышка, а затем распадается, образуя 1000 или больше отдельных ядрышек. Можно обнаружить до 3000 копий рДНК (образующих при центрифугировании четкую сателлитную область). Амплифицированная рДНК может служить удобным материалом для биохимических исследований. Так, например, структурные исследования, описанные в предыдущем разделе, были выполнены на ДНК именно этого типа.
Механизм амплификации ДНК интенсивно исследуется, но пока еще точно не установлен [283]. Было высказано предположение, согласно которому многочисленные копии рДНК образуются при помощи механизма разматывающего рулона, аналогично тому как это показано в уравнении (15-9). Значение амплификации рДНК состоит, вероятно, в том, что создаются условия для образования большого количества рибосом, необходимых для ускорения белкового синтеза.