Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Хромосома эукариот и ее контроль
Ядерные белки
В бактериальных клетках отрицательно заряженные фосфатные группы ДНК могут быть в значительной степени нейтрализованы положительно заряженными полиаминами. Однако основные белки также стремятся частично «одеть» ДНК. В зрелых головках сперматозоидов рыб плотно упакованная ДНК нейтрализуется протаминами — специальными низкомолекулярными белками (с мол. весом ~5000), богатыми остатками аргинина. Сходные основные белки обнаружены в сперме млекопитающих [284]. Однако в соматических клетках, отрицательные заряды ДНК компенсируются главным образом положительным зарядом гетерогенных групп основных белков, известных под названием гистонов. Существует пять классов гистонов, мол. вес которых составляет от ~11 000 до 21 500 [285—287]:
Н1 (или I, или f1)
Богатый лизином
Н2а (или IIb1, или f2a2)
Н2b (или IIb2, или f2b)
Умеренно богатые лизином
Н3 (или III, или f3)
Н4 (или IV, или f2а1)
Богатые аргинином
Отличительной чертой гистонов, богатых аргинином, является удивительное постоянство их аминокислотной последовательности. Так, гистон Н4 из пропростков гороха отличается от аналогичного гистона из тимуса крупного рогатого скота всего лишь двумя аминокислотами. Что же касается богатого лизином гистона Н1, то его последовательности почти присуща видовая специфичность.
Гистон Н3 из тимуса теленка содержит 135 аминокислотных остатков [288], причем суммарный заряд первых 53 из них составляет +18. Возможно, именно эта часть белка связывается с ДНК. В то же время карбоксильный конец этого гистона обладает гидрофобными свойствами и лишь в незначительной степени — основными. Интересные кластеры основных аминокислот были обнаружены в отдельных участках полипептидной цепи гистона Н2а [289]. Одна из любопытных особенностей строения гистонов — это наличие большого числа микромодификаций, сводящихся к фосфорилированию остатков серина, ацетилированию и метилированию остатков лизина, а также метилированию боковых цепей аргинина. Так, например, остатки Lys-14 и Lys-23 в гистоне Н3 N-ацетилированы, тогда как остатки Lys-9 и Lys-27 частично ε-N-метилированы — каждый участок содержит частично моно-, частично ди- и частично триметильные производные.
Какие же другие функции кроме нейтрализации зарядов ДНК выполняют гистоны? Первоначально считали, что эти белки могут играть роль репрессоров генов аналогично тому, как это происходит у бактерий. Однако экспериментального подтверждения это предположение не получило. Гистоны, по-видимому, образуют своеобразный комплекс с нитями ДНК. Сравнительно недавно с помощью электронного микроскопа были получены микрофотографии, на которых видно, что хроматиновые волокна имеют регулярно повторяющееся строение, напоминая нитки бус. Диаметр бусинки (или v-телец, или нуклеосом) составляет 7—10 нм, а длина свободной «нитки» между бусами равна 2—14 нм. (рис. 15-35] [290—294]. Содержание ДНК в «бусинках» велико. Данные, полученные методом дифракции нейтронов, свидетельствуют о том, что в у-частицах нить ДНК намотана вокруг гистонового олигомера, (рис. 15-36) [295]. Гистоны Н2а, Н2в, Н3 и Н4 обнаруживаются почти в одинаковом количестве — на каждые 100 пар оснований в ДНК приходится по одной молекуле каждого из гистонов. В растворе был получен октамер, содержащий по две субъединицы гистонов каждого типа [296].
При обработке нуклеазами хроматин быстро расщепляется на фрагменты, состоящие из 205+15 пар оснований, и более медленно — на фрагменты, состоящие из 170 пар оснований. Этот результат в сочетании с приведенными выше данными позволил предположить существование структуры, в которой фрагмент ДНК, состоящий из 200 пар оснований, обмотан вокруг гистонового октамера таким образом, что двухцепочечная нить ДНК длиной 68 нм упаковывается в одной у-частице размером порядка 10 нм. Соседние v-частицы связаны друг с другом очень короткими участками ДНК. Было высказано предположение, что обычная двойная спираль ДНК, поворачиваясь вокруг гистонов в v-частице, может претерпевать резкие «изломы» через каждые 20 пар-оснований [297], причем при каждом таком изломе спираль будет раскручиваться на 15—20°. Гистон Н1, присутствующий в меньшем количестве, чем другие гистоны, может играть роль агента, способствующего образованию поперечных связей в хроматине (рис. 15-35). Согласно другим данным [296а], на каждую v-частицу приходится один отрицательный виток суперспирали. Если это так, то число v-частиц на рис 15-35 соответствует числу витков суперспирали в ДНК вируса SV40 (рис. 2-27). Гистон Н1 взаимодействует преимущественно с суперспирализованной ДНК, [296b]. Исследования с использованием химических агентов, образующих поперечные связи, показали, что гистоны Н1 часто обнаруживаются в непосредственной близости от нескольких, других гистонов, причем гистон Н2В с равной вероятностью располагается рядом с Н2А, Н3 или Н4 [298]. Что касается хроматина животных, грибов и зеленых растений, то в основных чертах он имеет такое же строение [299].

РИС. 15-35. А. Электронная микрофотография «мини-хромосомы», образованной вирусом SV40, растущим в культивируемых клетках обезьяны [292].
В показанной нативной форме диаметр нуклеопротеидного волокна составляет приблизительно 11 нм, а длина — около 210 нм. Б. Мнни-хромосомы в виде нитки бус наблюдались при понижении ионной силы. Все 21 бусинки диаметром около 11 нм соединены друг с другом мостиками с диаметром, равным приблизительно 2 нм и длиной 13 нм. Опыты по депротеинизации и релаксации ДНК свидетельствуют о том, что общая длина показанной ДНК в семь раз превышает общую длину нативной мини- хромосомы. В. Электронная микрофотография хроматиды эмбриона Drosophila melanogaster бластодермальной стадии в процессе репликации. Обратите внимание на наличие нуклеосомных (у) частиц непосредственно вблизи репликационной вилки (McKnight. L., Miller О. L., Jr.).
На основе описанных выше данных была сформулирована современная точка зрения, согласно которой основная функция гистонов состоит в том, чтобы обеспечить необходимую упаковку ДНК. Однако иногда гистон Н1 называют общим репрессором, удерживающим хроматин в компактно упакованном состоянии, препятствующем транскрипции. Поскольку процесс инициации митоза сопровождается фосфорилированием гистона Н1 при помощи специальной протеинкиназы, можно предположить, что этот гистон играет какую-то иную роль [300]. Другие гистоны, особенно F4, подвергаются множеству модифицирующих воздействий, в том числе ацетилированию и фосфорилированию (обратимо) и метилированию (необратимо) [301]. Значение этих реакций в регуляции таких процессов, как транкрипция и репликация, до сих пор неясно.
Какие еще белки кроме гистонов обнаруживаются в клеточных ядрах? Методом электрофореза в полиакриламидном геле было установлено, что в ядрах клеток HeLa содержится около 450 компонентов, большинство из которых присутствует в небольших количествах (<10000 молекул в расчете на одну клетку) и не обнаруживается в цитоплазме [302]. К наиболее кислым белкам относится большое число ферментов, включая РНК-полимeразу. Кроме того, в ядрах содержатся: 1) определенные репрессоры генов, в основном не идентифицированные, 2) белки, связывающие гормоны, и 3) многие другие белки [303]. Наряду с ядерными белками, которым уделяется обычно основное внимание, определенную роль в регуляции фенотипического выражения генов играет также мало исследованный класс небольших ядерных РНК. Молекулы этой РНК длиной от 65 до 200 нуклеотидов могут стимулировать транскрипцию специфических генов, связываясь с комплементарными участками ДНК. Таким образом, информация, транскрибированная с одного участка хромосомы, может оказывать влияние на процессы, протекающие на другом участке или на другой хромосоме [303а].
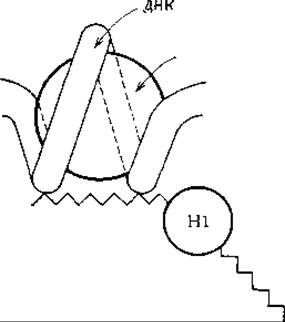
РИС. 15-36. Схематическое изображение предполагаемого строения субъединицы хроматина [295].
Белковый стержень представляет собой комплекс аполярных сегментов четырех указанных в тексте гистонов. Участки гистонов, обладающие основными свойствами, образуют комплекс с ДНК, располагающейся на поверхности нуклеосомы. Гистон Н1, расположенный между нуклеосомами, может играть роль агента, образующего поперечные связи либо между нуклеосомами одной и той же цепи, либо между нуклеосомами разных цепей. Шаг спирали ДНК не обязательно должен быть постоянным; при среднем диаметре нуклеосомы около 10 нм он равен 5,5 нм.