Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Рост, дифференцировка и химическая коммуникация клеток
Дифференцировка тканей и биология развития
Изменения в содержании ДНК
Известно много случаев, когда дифференцировка клеток приводит к временному или устойчивому изменению генома. Например, амплификация генов рРНК (гл. 15, разд И, 1, д) ооцитов сопровождается временным увеличением общего содержания ДНК в клетке. Некоторые высокоспециализированные клетки, например клетки Пуркинье в мозжечке или многие клетки в личинках двукрылых (гл. 15, разд. Г. 9, в), характеризуются полиплоидией. Как правило, это клетки, достигшие предельного уровня специализацин и утратившие способность к делению. В каждой копни ДНК полиплоидных клеток обычно содержится полный набор генов (большая часть которых фенотипически не выражается).
С другой стороны, в некоторых клетках процесс необратимой дифференцировки сопряжен с потерей части генома. Крайним выражением этой ситуации являются эритроциты человека, полностью утратившие ядро. В других клетках разрушаются отдельные хромосомы. Возможны и такие случаи, когда хромосома или ее часть необратимо инактивируется и остается в клетке в виде компактного образования — гетерохроматина. Этим термином обозначают интенсивно окрашивающиеся области клеточного ядра. Некоторые гетерохроматины содержат многократно повторяющиеся последовательности (гл. 15, разд. И, 1,6), но в отдельных гетерохроматиновых областях обнаруживаются группы инактивированных генов. Чрезвычайно интересен случай полной инактивации одной из двух Х-хромосом в клетках самок млекопитающих [181]. Вся хромосома при этом выглядит как гетерохроматин. Инактивация происходит на ранней стадии эмбрионального развития и захватывает ту или другую Х-хромосому по принципу случайности: в одних клетках инактивируется материнская Х-хромосома, в других — отцовская. Однако при дальнейших клеточных делениях одна и та же хромосома остается инактивированной во всем клоне клеток. В результате в организме особей женского пола возникает мозаицизм по гетерозиготным генам Х-хромосом.
Механизм инактивации и избирательного разрушения хромосом остается неизвестным; предполагается, что в своей химической основе он имеет сходство с процессами модификации и рестрикции, свойственными бактериям (гл. 15, разд. Е) [182]. С помощью системы метилирования одна хромосома может быть «помечена» и сохранена, тогда как другая — подвергнуться последовательным воздействиям отстригающей эндонуклеазы. С другой стороны, не исключено, что какой-то другой фермент инициирует переход хромосом в гетерохроматин.
Если одни гены избирательно инактивируются или попеременно включаются и выключаются, то другие в некоторых случаях необратимо утрачиваются в процессе клеточной дифференцировки. В хромосомах отдельных клеток во время митоза, по-видимому, имеет место генетическая рекомбинация. Был обнаружен кроссинговер между сестринскими хроматидами. Однако если при этом происходит обмен равными количествами генетического материала, то изменения генетики дочерних клеток не наступает. С другой стороны, если в одной молекуле ДНК оказываются две и более одинаковые последовательности оснований, то возможен неравный кроссинговер (гл. 16, разд. Ж, 3) с потерей генетического материала одной из дочерних клеток. По существу в этом может состоять предопределенная программа дифференциации для некоторых клеток.
Потеря генов из хромосом может идти и по Другому механизму, а именно путем исключения петли ДНК (looping оut mechanism) [183].
По аналогии с исключением профага А, (гл. 15, разд. Г, 8) из хромосомы Е. coli такая потеря генов должна происходить в специфических Сайтах (участках) ДНК. Постоянная потеря генетического материала может, по-видимому, происходить при дифференцировке плюрипотентных стволовых клеток, образующих клетки крови. Из указанных плюрипотентных клеток сначала формируются три другие линии стволовых клеток, а именно, миелоидные, эритроидные и лимфоидные, которые подвергаются дальнейшей дифференцировке, как показано на схеме.
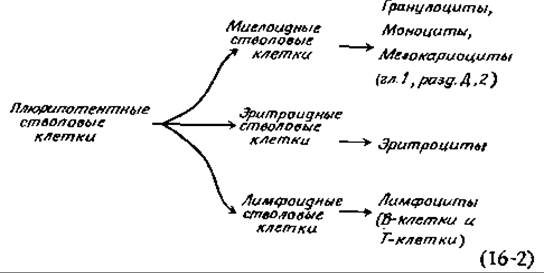
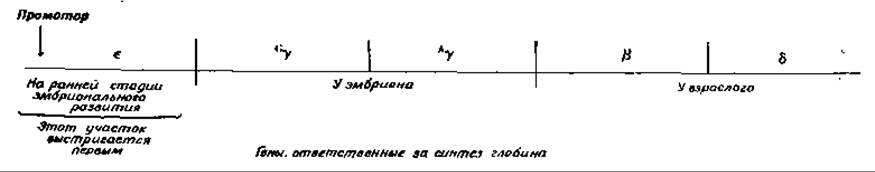
Эритроидные стволовые клетки служат предшественниками содержащих гемоглобин эритроцитов. Вспомним (гл. 4, разд. Д, 7), что гемоглобины млекопитающих состоят из двух a-цепей и еще двух других цепей — либо ß, либо у, либо б, либо є. Гемоглобин взрослых в основном имеет структуру a2ß2, но имеется также небольшое количество гемоглобина а2δ2. Для эмбриона на ранних стадиях развития характерен гемоглобин а2ε2, но на последующих стадиях ε-цепи замещаются двумя другими, свойственными эмбриональному гемоглобину цепями, а именно Gу и Ay. Генетические исследования показали, что гены e-, y-, ß- и 6-глобина тесно сцеплены [188]. Почему же в отдельном эритроците присутствует гемоглобин только одного типа? Видимо, дело в том, что для данного набора генов существует только один промотор. Если после каждого гена имеется сигнал-терминатор, то очевидно, что будет идти транскрипция только того гена, который ближе всех прилегает к промотору. В случае потери на каком-то этапе развития этого гена начнет транскрибироваться следующий ген и т. д.; таким образом могут происходить нарастающие постепенные изменения в выражении гена в эритроцитах. Еще одна особенность процесса дифференцировки эритроцитов — это его чувствительность к гормону эритропоэтину, гликопротеидному гормону, образующемуся в почках [184—186]. Под действием эритропоэтина в дифференцирующих стволовых клетках начинается интенсивный синтез гемоглобина, и они окончательно превращаются в эритроциты [186а].