Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Свет в биологии
Зрение
Строение наружного членика палочек
Фоторецепторы глаза выполняют совершенно иные функции, чем хлоропласты. Зрительные рецепторы предназначены для инициации нервного импульса, и поэтому их главным свойством является высокая чувствительность — некоторые рецепторы улавливают практически каждый падающий на них фотон. Этой цели служат многослойные мембраны, в которые включены в большом количестве сильно поглощающие молекулы [133, 133а].
Сетчатка человеческого глаза содержит свыше 108 плотно упакованных рецепторных клеток двух типов — палочек и колбочек. Палочки представляют собой чрезвычайно чувствительные клетки, способные реагировать всего на пять квантов света. Предназначенные для зрения в условиях слабой освещенности, они дают «черно-белую картину» и сконцентрированы на периферий сетчатки. Менее чувствительные колбочки расположены в основном в центре сетчатки и делятся на три типа, различающиеся по спектральной чувствительности. Они обеспечивают цветовое зрение.
Для палочек (рис. 13-28), исследованных более детально, чем другие рецепторы сетчатки, характерен весьма интенсивный метаболизм. Палочки человеческого глаза могут функционировать в течение сотни лет [134]. В них наблюдается замечательный процесс самообновления, при котором старые мембранные диски на конце палочки отбрасываются [135] и заменяются новыми со стороны, ближайшей к ядру. Наружный членик окружен плазматической мембраной. Внутри мембраны (по-видимому, без присоединения к ней) находятся ~500 параллельно уложенных дисков диаметром ~2 мкм, расстояние между центрами которых составляет ~32 нм. Каждый диск образован парой мембран толщиной ~7 нм, разделенных очень узким пространством. На электронных микрофотографиях пространство внутри дисков как бы замкнуто по краям. Соседние диски разделены несколько большим пространством.
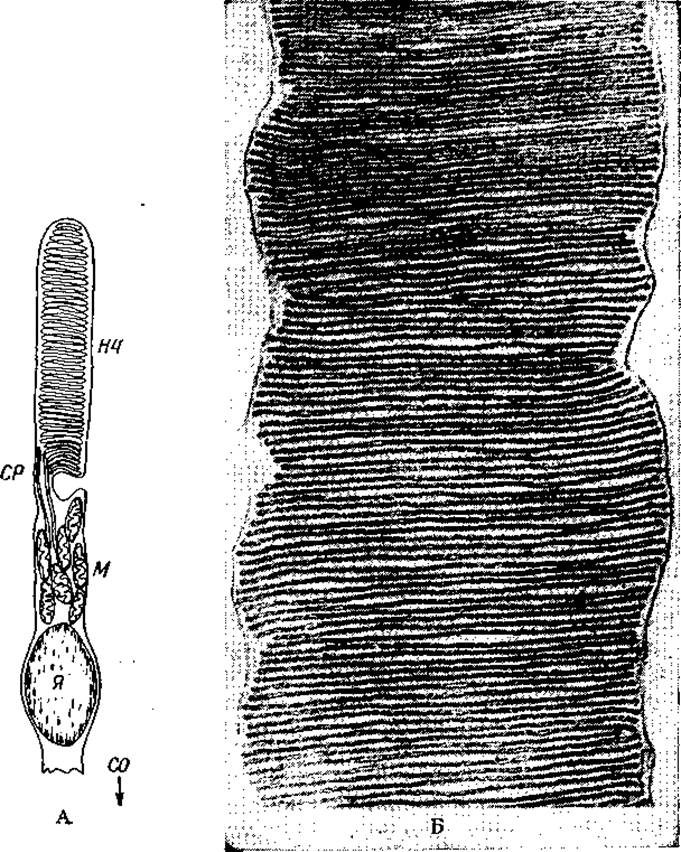
РИС. 13-28. А. Схема строения палочки сетчатки позвоночных [135]. НЧ — наружный членик; СР — соединительная ресничка; М — плотно упакованные митохондрии; Я — ядро; СО — синаптическое окончание. Б. Электронная микрофотография продольного среза наружного членика палочки в сетчатке крысы (с любезного разрешения Дж. Даулинга [133b]).
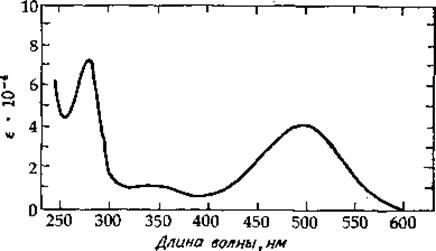
РИС. 13-29. Спектр поглощения родопсина быка, диспергированного в воде в присутствии неионного детергента. [Shich Н. et al., JBC, 244, 529—536 (1969).]
Мембраны дисков на 60% состоят из белка и на 40% — из липида (табл. 5-1). Около 80% белка приходится на долю родопсина (зрительный пурпур) — липопротеида, нерастворимого в воде, но растворимого в растворах детергентов. Для диспергирования молекул родопсина широко используется дигитонин, не меняющий оптических свойств родопсина. Родопсин млекопитающих имеет мол. вес ~ 28 000—35 000; каждая молекула содержит один хромофор, характеризующийся λmах — 500 нм (рис. 13-29). Поскольку родопсин составляет основную Массу мембранного белка, очевидно, что его молекулы тесно примыкают одна к другой. На срезе палочек, приготовленном методом замораживания — травления, выявляются круглые частицы диаметром ~4—5 нм, слишком крупные, чтобы их можно было считать отдельными молекулами родопсина (если только каждая молекула не окружена слоем липида). Эксперименты By и Страйера по переносу энергии (разд. В, 2) привели к заключению, что молекула родопсина имеет Удлиненную форму (с максимальным размером 7,2 нм) и может пронизывать мембрану. Было высказано предположение, что эта молекула имеет крупную, почти сферическую головку, расположенную с внутренней стороны мембраны, и тонкий хвост, пронизывающий мембрану [136]. Хромофор расположен так, что вектор дипольного момента перехода оказывается параллельным плоскости дисков (т. е. перпендикулярным направлению движения падающих фотонов).