Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Метаболизм азотсодержащих соединений
Соединения, образующиеся из аспартата
Метаболизм метионина
Метионин включается в белки и как таковой, и в виде N-формилметионина в качестве N-концевого остатка бактериальных белков (рис. 14-9, стадии а и б). Как в клетках животных, так и в клетках растений метионин может подвергаться переаминированию в соответствующую-кетокислоту (стадия в), но в количественном отношении эта реакция едва ли имеет важное значение. Главный путь превращения метионина связан с его превращением в S-аденозилметионин (SAM, рис. 14-9, стадия г). Эта реакция уже обсуждалась (гл. 11, разд. Б, 2); была рассмотрена (гл. 7, разд. В, 2) и функция SAM в процессе трансметилирования (стадия д). Продукт трансметилирования S-аденозилгомоцистеин превращается в гомоцистеин путем необычной гидролитической реакции отщепления аденозина (стадия е)1). Гомоцистеин может быть, снова превращен в метионин, как показано штриховой линией на рис. 14-9, а также в уравнении (8-85). Другой важный путь метаболизма гомоцистеина связан с превращением в цистеин (рис. 14-9, стадии ж и з). Эта последовательность реакций обсуждается в разд. Ж. Другим продуктом на этом пути является а-кетобутират, который доступен окислительному декарбоксилированию с образованием пропионил-СоА и его дальнейшим метаболизмом или может превращаться в изолейцин (рис. 14-10).
В растениях SAM может быть использован в качестве субстрата в интересной реакции, приводящей к образованию этилена [59, 60]. Этилен, как было установлено еще в 1858 г., вызывает у растений утолщение стебля и снижает скорость его удлинения. В 1917 г. было показано что это соединение образуется в плодах и что при добавлении этилена в газовую среду скорость созревания плодов повышается. В настоящее время твердо установлено, что этилен относится к гормонам растений1, он вызывает множество эффектов, включая замедление митоза. На синтез этилена отчасти влияет гормон ауксин (гл. 16, разд. А, 3), а также красный свет. Известно, что кетокислотный аналог метионина в присутствии Н2О2 подвергается окислительному декарбоксилированию с образованием этилена и двух молекул СО2. Однако в растениях СО2 образуется из атома С-1 метионина, а С-2 переходит в формиат (рис. 14-9, стадия и). Кроме того, —S—СН3-группа каким-то образом вновь поступает в метионин. Если считать, что предшественником этилена является SAM, то другим продуктом должен быть, очевидно, 5'-тиометиладенозин, как это показано на рис. 14-9. Последнее соединение образуется в организме животных в ходе важной реакции катаболизма SAM (рис. 14-9, стадия к). Реакция представляет собой внутримолекулярное замещение у-метиленовой группы SAM карбоксилатной группой.
1) Было показано, что фермент содержит прочно связанный NAD+. Вероятно, он используется для окисления аденозина по 3'-положению, что и приводит к элиминированию гомоцистеина. Присоединение воды и восстановление дают аденозин [58а).
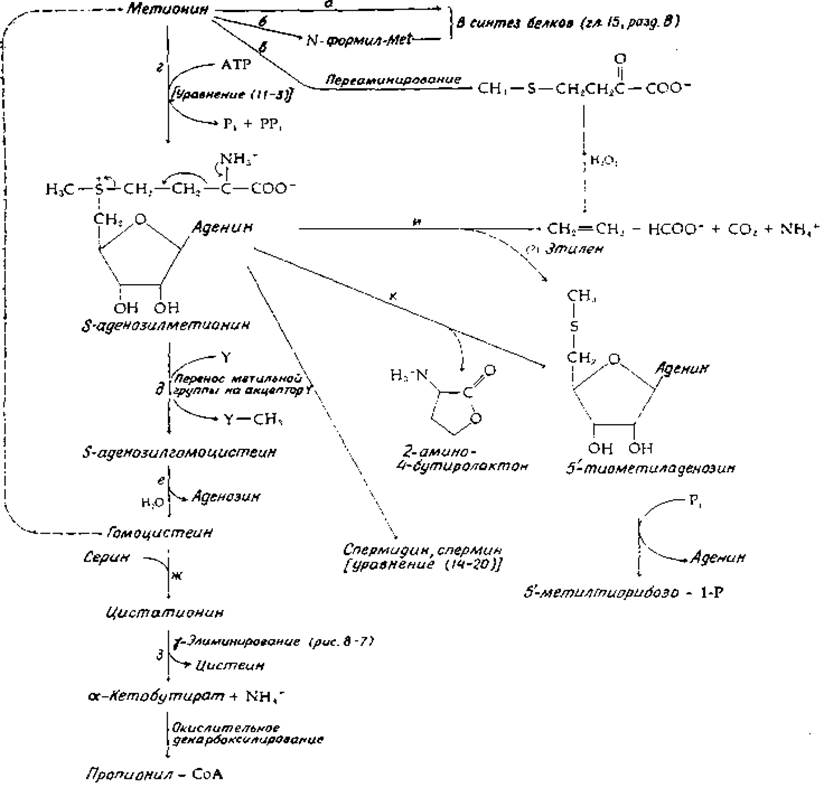
РИС. 14.9. Некоторые реакции метаболизма метионина.
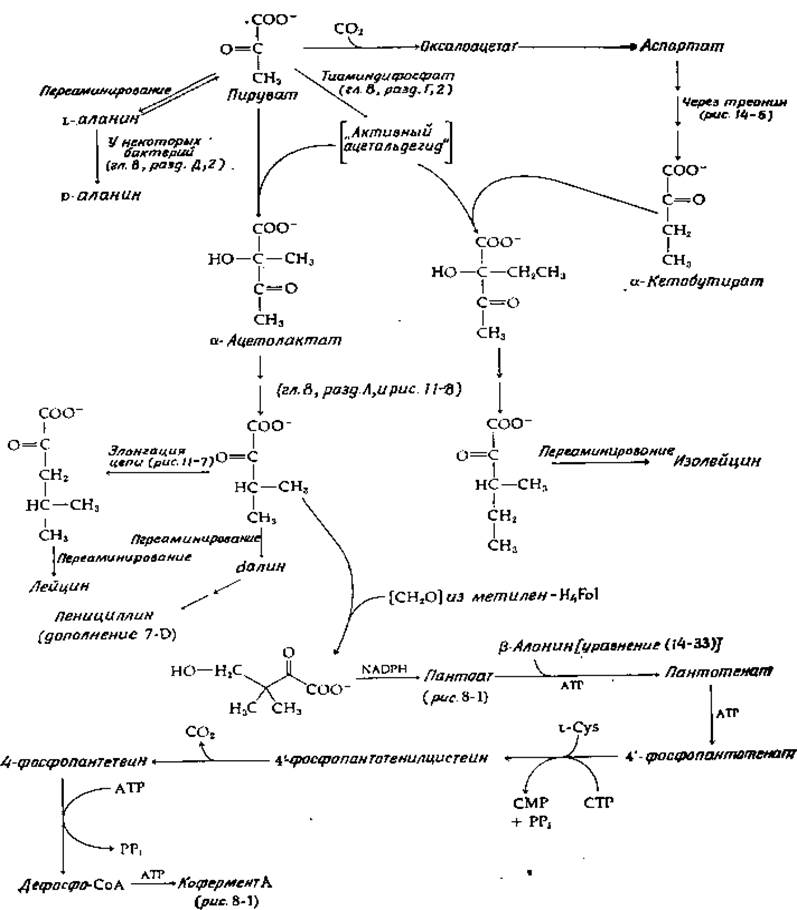
РИС. 14-10. Биосинтез лейцина, изолейцина, валина и кофермента А.