Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Транскрипция молекул РНК
Транскрипция в эукариотических клетках
В клетках с истинным ограниченным мембраной ядром молекулу информационной РНК живут сравнительно долго. За время своей жизни они должны выйти из ядра в цитоплазму, к местам синтеза белка. Помимо очевидной необходимости жить дольше и перемещаться на большие расстояния по сравнению с бактериальным мРНК эукариотические мРНК отличаются от них еще по целому ряду до конца не изученных параметров. Вполне возможно, например, что в случае эукариот молекулы мРНК транскрибируются с отдельных генов и полигенные опероны у них обычно не функционируют. В эукариотических клетках был обнаружен новый тип РНК, получивший название гетерогенной ядерной РНК (Г-яРНК). Существует предположение, что эта РНК, на долю которой приходится основная часть ядерной РНК, служит предшественником мРНК. Подобно мРНК эта РНК похожа по нуклеотидному составу на ДНК. Мол. вес этой РНК лежит обычно в пределах от 105 до 2∙107 (что соответствует содержанию 1500—30 000 нуклеотидов). Для молекул Г-яРНК характерна способность к быстрым превращениям — для большинства из них время полужизии не превышает 10 мин. Однако некоторые из Г-яРНК могут существовать и 20 ч. Вызывает удивление тот факт, что из ядра выходит лишь около 10% Г-яРНК, а большая ее часть распадается в ядре, так и не переходя в цитоплазму [77—79].
Неожиданное открытие помогло подтвердить, что Г-яРНК действительно является предшественником мРНК. Было показано, что как у Г-яРНК, так и у мРНК эукариот на 3'-конце находятся длинные цепи полиадениловой кислоты [поли(А)]. (Исключение, по-видимому, составляют молекулы мРНК, кодирующие синтез гистонов — основных ядерных белков, обнаруживаемых в эукариотических клетках.) Обычно к концу цепи Г-яРНК присоединяется 200 остатков адениловой кислоты — по-видимому, при помощи, специфического фермента, действующего уже после завершения процесса транскрипции [79а].
Имеются данные о том, что поступающая в цитоплазму мРНК, образовавшаяся в результате расщепления Г-яРНК, все еще содержит 50—75 остатков адениловой кислоты. Эти остатки постепенно (однако, по-видимому, не полностью) отщепляются от мРНК. Какова функция таких участков поли (А)? Мы не знаем ответа на этот вопрос. Согласно одному из предположений, поли (А) зачем-то нужны для транспорта мРНК из ядра. Существует и другое предположение, а именно что после каждого акта трансляции, когда мРНК покидает рибосому, от ее 3'-конца отщепляется один или несколько нуклеотидов. Таким образом, согласно этому объяснению, отдельные звенья 3'-концевых участков поли (А) — это как бы «билеты», каждый из которых дает ей право на одну «поездку» через рибосому. Состояние, когда все остатки А уже израсходованы, служит клетке сигналом для разрушения мРНК. Хотя считается, что наличие участков поли (А) в мРНК является характерной особенностью эукариотических клеток, быстро распадающиеся последовательности поли (А) обнаруживаются также и у Е. coli [80].
Другая удивительная особенность мРНК, которая была обнаружена сравнительно недавно, состоит в том, что, как у эукариот, так и у вирусов 5'-концы молекул многих мРНК прикрыты концевыми «колпачками», имеющими специфическую структуру и содержащими7-метилгуанозин в виде биполярного иона, образующегося в результате отщепления протона [81—83]. Обратите внимание на трифосфатный мостик, при помощи которого 7-метиладенозин соединен с мРНК. Он может образовываться следующим способом [82]. На 5'-конце РНК-транскрипта первоначально находится трифосфатная группа, поскольку при инициации транскрипции затравкой служит нуклеозидтрифосфат. Одна из таких фосфатных групп может отщепляться, после чего остается дифосфат, который и взаимодействует с GTP. После этого 7-метильная группа, а также 2'-метальная группа рибозы1), находящейся на 5'-конце полинуклеотида, переносится с S-аденозилметионина при помощи соответствующей метилазы.

мРНК полиовируса необычна в том отношении, что в инфицированных клетках она остается неприкрытой концевым колпачком [83а].
Ядрышко
В результате многочисленных исследований было окончательно установлено, что местом синтеза рибосомной РНК является ядрышко (в ядре может быть одно или несколько ядрышек). Рибосомы эукариотических клеток содержат четыре различных типа молекул РНК, размеры которых указаны ниже.
1) Следующий остаток рибозы также иногда метилируется, и N6-метиладенозин может находиться на 3'-конце рядом с участком поли (А) [83].
|
Константа седиментации (S) |
Мол. вес |
число нуклеотидов |
|
28 |
1,7∙106 |
5000 |
|
18 |
0,65∙106 |
2000 |
|
5,8 |
5∙104 |
150 |
|
5 |
4∙104 |
120 |
Молекулы 28S, 18S и 5,8S образуются из высокомолекулярной (4∙106) 45S-пpe-pPHK. Этот предшественник транскрибируется в центральной зоне ядрышка. При выходе молекулы пре-рРНК в наружную «кору» она поэтапно распадается [уравнение (15-6)] [57, 84, 85].

Методом электронной микроскопии было прямо подтверждено наличие взаимосвязи одной молекулы предшественника с другой (рис. 15-11) [85а, b]. Заметим, что ближе других к 5'-концу находится 18S-субъединица 45S-PHK, точно так же, как 16S-pPHK в большом транскрипте генов рРНК прокариот (разд. Б,3). Возможно, что число этапов в действительности больше, чем показано на рисунке. Так, образованию 45S-PHK может предшествовать образование 46S- и 47S-молекул.

РИС. 15-11. А. Электронная микрофотография 45S-предшественника рРНК клеток Hela после «распрямления» молекул в 80%-ном растворе формамида и 4M мочевине. Изображение молекулы получено методом негативного контрастирования. Б. Контуры молекулы, изображенной на рис. А. Видны несколько участков, обладающих вторичной структурой двухцепочечных шпилек. Показаны участки, соответствующие 28S- и 18S-pPHK. В. 32S-pPHK. Г. 28S-pPHK. Обратите внимание, что в 28S-pPHK можно видеть те же вторичные структуры, что и в ее 32S- и 45S-предшественниках [85а].
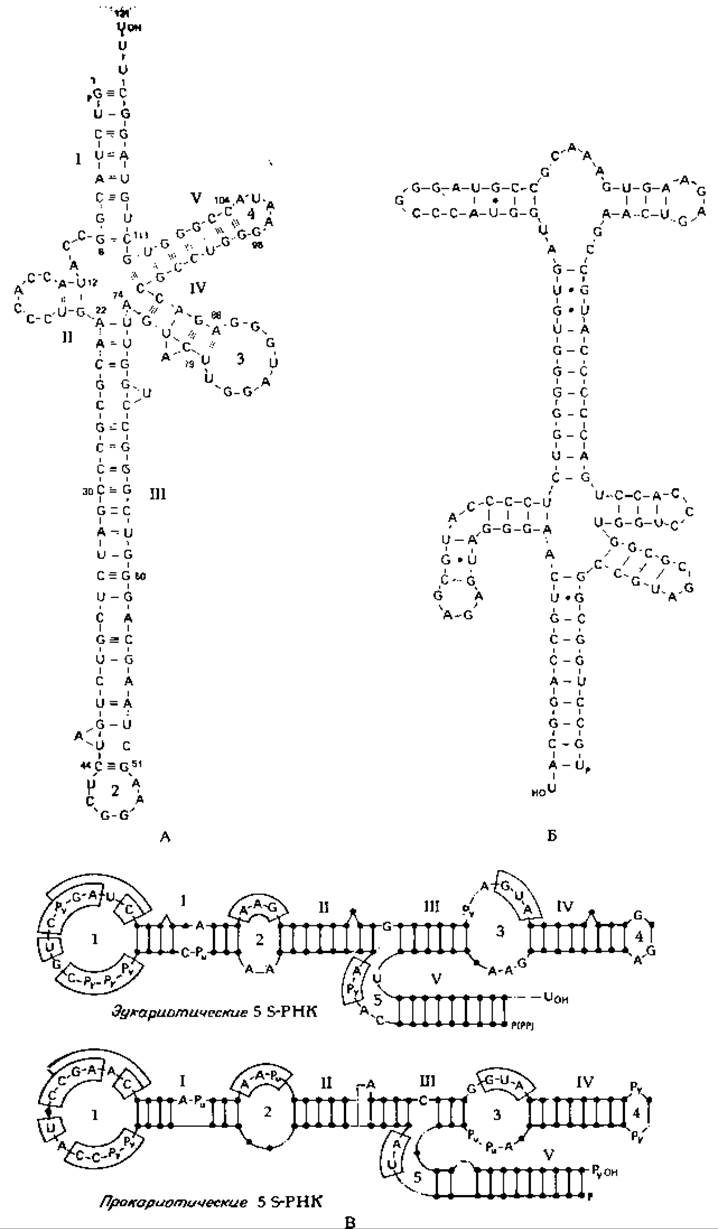
РИС. 15-12. Нуклеотидная последовательность рибосомной 5S-PHK клеток человека (KB-карцинома). А и Б. Две возможные структурные формы (Sirlin J. L. «Biology of RNA», p. 67, Academic Press, New York, 1973). В. Возможные варианты спаривания оснований в молекулах, 5S-PHK эукариот (сверху) и 5S-PHK прокариот снизу. Буквами внутри петель обозначены общие нуклеозиды для 5S-PHK KB-клеток человека, X. laevis и T. utilis (сверху) и для 5S-PHK E. coli, Р. fluorescens и В. stearothermophilus (внизу). Нуклеотиды, обведенные в рамки, обнаруживаются в указанных местах 5S-PHK как эукариот, так и прокариот (Nishikawa К., Takemura S., J. Biochem., Tokyo, 76. 935—947. 1974).
Необычайно важную роль в исследовании ядрышка сыграло прямое наблюдение неупорядоченной центральной зоны ядрышек при помощи электронного микроскопа [59, 86]. На ДНК-цепях пре-РНК-генов удалось увидеть образующиеся нити РНК, покрытые белком, (рис. 15-7). С одного гена одновременно транскрибируется приблизительно 80—100 РНК-цепей разной длины. Общая длина гена, согласно электронно-микроскопическим данным, составляет 2,3 мкм, что лишь ненамного меньше рассчитанной длины полностью вытянутой молекулы ДНК (в В-форме). Однако, судя по длине образующихся транскриптов, цепи пре-рРНК многократно сложены с образованием компактной структуры.
У любых организмов в гаплоидном наборе генов содержится, как правило, лишь один ген данного типа. Что же касается рибосомной РНК, то ее гены представлены в одном геноме множеством копий. Так, например, у дрозофилы обнаруживается 130—190 копий гена 45S-PPHK.
Ген рибосомной 5S-PHK у эукариот не связан с геном 45S-PHK и локализован не в ядрышке. У дрозофилы около 500 копий гена 5S-PHK расположены в правом плече хромосомы 2. За синтез 5S-PHK (нуклеотидная последовательность которой показана на рис. 15-12) ответственна РНК-полимераза III. Характерная особенность 5S-PHK состоит в том, что она может быть сложена по-разному, и до сих пор не ясно, каким способом или способами она укладывается в рибосомах [87].