Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Биохимическая генетика и синтез нуклеиновых кислот и белков
Генетические методы
Условно-летальные мутации
Изучение ауксотрофов по питательным веществам сыграло важную роль в развитии биохимии, но, к сожалению, с помощью этого метода можно изучать только один ген или группу генов, участвующих в синтезе какого-либо конкретного субстрата. Было бы желательно, однако, располагать методами обнаружения мутаций всего набора генов, содержащихся в клетках. Но этому препятствовало то обстоятельство, что большинство мутаций детальны, причем во многих случаях устранить этот эффект добавлением всевозможных субстратов не удается. Ранние генетические исследования показали, что летальные мутации особенно широко распространены среди высших организмов. Однако поскольку в эукариотических клетках существуют пары гомологичных хромосом, летальные мутации в одной из хромосом могут и не приводить к гибели организма. У бактерий и вирусов есть только одна хромосома, в связи с чем организмы с летальными мутациями выжить не могут.
Ауксотрофы по питательным веществам можно назвать условнолетальными мутантами. Это означает, что они могут выжить лишь при условии, что в среде имеется субстрат, синтез которого катализируется утерянным ферментом. Были обнаружены другие типы условно-летальных мутаций, позволяющие изучать почти любой ген в организме. Один класс таких мутаций называют температурочувствительными [135, 136]. Температурочувствительные (ts — от англ. temperature sensitive) мутанты отлично растут при низкой температуре, например при 25 °С, но не растут при более высокой температуре, например при 42 °С. При многих температурочувствительных мутациях имеет место такая замена аминокислот, в результате которой вновь образованный дефектный белок оказывается менее устойчивым к нагреванию, чем белок дикого типа. При других мутациях теряется способность к синтезу белков по причинам, которые не всегда ясны. Многие температурочувствительные мутации, несомненно, встречаются в природе. В качестве примера можно указать на ген, контролирующий окраску волос у сиамских кошек [136]. Ген (или его продукт) неактивен при температуре тела, но активен в плохо обогреваемых частях тела —в лапках, в хвосте и в области носа. В результате шерсть кошки окрашена лишь в этих участках.
В результате поиска условно-летальных температурочувствительных мутантов бактериофага Т4 были обнаружены сотни различных мутантов с беспорядочным распределением мутаций по всей длине фаговой хромосомы. С помощью комплементационного анализа были выявлены отдельные соответствующие им гены, для которых раньше было установлено только общее число (рис. 15-21). В дальнейшем были определены специфические функции многих из этих генов [137, 138]. Так, оказалось, что продукт гена 42 представляет собой специфический фермент, необходимый для синтеза оксиметил-dCMP (гл. 14, разд. Л,1). Гены 20—24 в числе прочих, по-видимому, кодируют синтез белков головки фага, так как при мутациях этих генов образуются нормальные отростки, но белки головки не синтезируются. Ген 23 кодирует, по-видимому, синтез основных субъединиц головки, а ген 20 как-то связан с процессом образования «колпачка» (capping) на конце головки. Эти мутанты образуют цилиндрические «полиголовки» вместо обычных головок. У мутантов с измененными генами 25—29 нарушено строение базальной пластинки и совсем не образуется отросток, а у мутантов с измененными генами 34—38 отсутствуют нити отростка.
Второй тип условно-летальных мутаций связан с превращением какого-нибудь из аминокислотных кодонов в один из трех кодонов терминации полипептидной цепи UAG, UAA и UGA (табл. 15-2) [139, 140]. Эти мутации часто называют nonsense-мутациями в отличие от missense-мутаций, при которых происходят аминокислотные замены. У мутанта с преждевременным окончанием синтеза полипептидной цепи синтезируется только часть продукта дефектного гена, после чего из-за наличия терминирующего кодона полипептидная цепь освобождается с рибосом. Интересная особенность мутаций с терминацией цепи заключается в том, что они могут быть супрессированы другими мутациями, локализованными в более отдаленных участках хромосомы вируса или бактерии. Так, например, многие условно-летальные мутации бактериофага Т4 были обнаружены благодаря их способности расти в определенных мутантных штаммах Е. coli (содержащих супрессорные гены) [140а] и неспособности расти в нормальном штамме В. Три различных супрессорных гена supD, supE и supF, для которых используют также обозначения su1, su2 и su31) супрессируют мутации, приводящие к появлению кодона UAG. Гены этого типа называют обычно аmbеr-супрессорными генами (см. ссылку на стр. 194). Были обнаружены и другие супрессоры этого класса. Другой класс мутаций, вызывающих преждевременную терминацию цепи, супрессируется генами supB и supC и включает мутацию к UAA. Обычно их называют ochre-мутациями. Сравнительно недавно были найдены супрессоры мутаций, приводящих к появлению кодона UGA [140в].
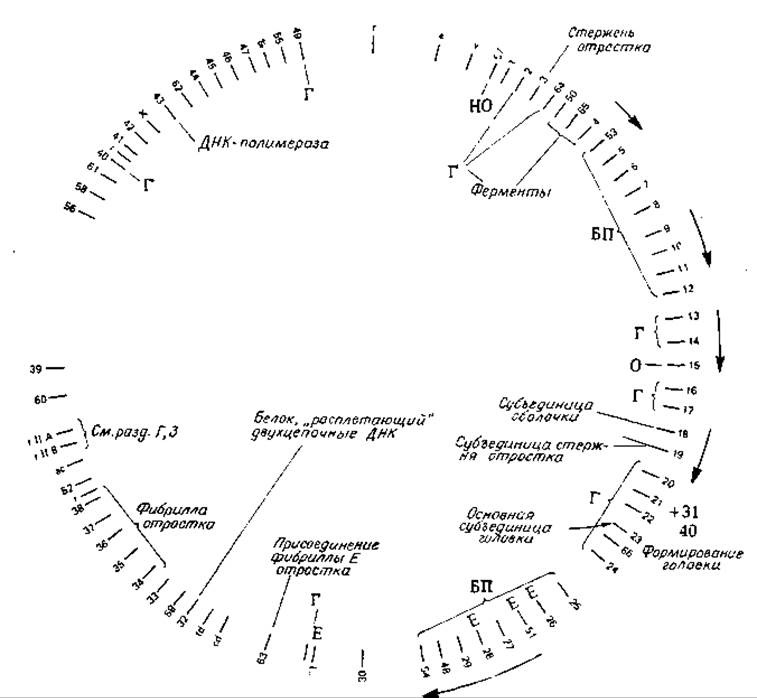
РИС. 15-21. Генетическая карта бактериофага Т4 (Sober Н. А., ed., CRC. Handbook of Biochemistry, 1st ed., p. 1—25, Chem. Rubber Publ. Co., Cleveland, Ohio, 1968, and Eiserling and Dickson, Annu. Rev. Biochem., 41, 467—502 (1972).
Сокращения: Г — головка, О — отросток, НО — нити отростка, БП — базальная пластинка, Е — белки, у которых ферментативная функция выражена сильнее, чем структурная. Стрелки, идущие вдоль окружности, охватывают группы совместно транскрибируемых генов.
Подобно температурочувствительным мутантам, amber- и ochre-мутанты могут быть получены практически для любого гена бактериофага. Мутантов с терминацией цепи, у которых утерянные гены не имеют жизненно важного значения для бактерий, можно распознать, обеспечив перенос генов либо путем скрещивания, либо путем вирусной трансдукцин в штамм (sup+), содержащий необходимый супрессорный ген.
1) Обозначения sup D, sup Е и sup F были предложены Бахманом и др. [15].
Условно-летальные мутанты сыграли чрезвычайно важную роль в изучении генетики бактериальных вирусов. Они были использованы также в качестве мощного метода при изучении сложных проблем, связанных с физиологией бактерий. Так, например, насколько сложно устроена система, необходимая бактерии для того, чтобы почувствовать наличие в среде питательного вещества и «подплыть» к нему? Оказалось, что бактерии «запрограммированы» чувствовать градиенты концентраций химических аттрактантов и менять направление движения таким образом, чтобы оказываться в области с более высокой концентрацией [141, 143]. Было бы интересно узнать, какое количество белков необходимо для того, чтобы чувствовать аттрактант, передавать необходимый информационный сигнал жгутикам (дополнение 4-Б) и направлять движение последних, вызывая их вращение, приводящее либо к передвижению вперед, либо к беспорядочному подергиванию (гл. 16, разд. Б,7).
Несмотря на то что сейчас выяснены лишь некоторые ключевые моменты тех химических процессов, которые лежат в основе всех этих явлений, использование температурочувствительных мутантов и тестов на комплементацию поможет установить суммарное число генов, принимающих участие в этих процессах, а также локализацию этих генов в хромосоме Е. coli. В ряде случаев это может способствовать более полному пониманию биологического явления.