Принципы структурной организации белков - Г. Шульц 1982
Эволюция белков
Обнаружение отдаленных родственных связей
Сравнения укладки цепи
Укладка цепи выявляет отдаленные эволюционные родственные связи. В предыдущих разделах упоминалось о том, что в процессе эволюции характер свертывания полипептидной цепи в пространстве сохраняется значительно лучше, чем ее аминокислотный состав. Поэтому тип свертывания цепи позволяет устанавливать такие отдаленные родственные связи, которые уже не проявляются в аминокислотных последовательностях. Для этого необходимо количественное описание способов свертывания цепей и сравнений укладки цепей.
Средние Са-расстояния служат обычным критерием подобия укладки цепей. Тип свертывания цепи хорошо описывается координатами Са-атомов. Поэтому укладку цепей можно сравнить путем наложения двух цепей с помощью минимизации расстояний между соответствующими Са-атомами [600]. Среднеквадратичное или обычное среднее значение полученных расстояний между соответствующими Са-атомами можно принять за показатель подобия при сравнении. Такой показатель (среднеквадратичное Са-расстояние) в настоящее время является общепринятым. Его применяют, например, для проверки результатов моделирования свертывания цепи (разд. 8.6). Были также предложены и использованы для сравнения свертывания цепей [601] и более сложные показатели подобия, основывающиеся на расстояниях Са, однако эти показатели не получили большого распространения.
Стандартную значимость для сравнения типов свертывания цепей пока определить нельзя. При сравнении свертывания цепей можно использовать среднеквадратичное Са-расстояние таким же образом, как использовалась сумма М, введенная для сравнений аминокислотных последовательностей. Однако если кумулятивная вероятность для М могла быть рассчитана путем сравнения со случайными аминокислотными последовательностями (рис. 9.5, а), то в случае свертывания цепей ситуация более сложна, поскольку случайные укладки цепи неизвестны. Поэтому пока еще невозможно установить стандартную значимость такого сравнения, как это было сделано при сопоставлении последовательностей миоглобина и а-цепи гемоглобина, приведенном на рис. 9.5, а.
Генерация и сравнение случайных свертываний цепи для нахождения кумулятивных вероятностей среднеквадратичных Са-расстояний требует больших объемов расчета [387]. Кроме того, при расчете таких распределений следует установить различия между структурным сходством, вызванным физико-химическими ограничениями (т. е. сходством вторичных и сверхвторичных структур) и сходством свободно варьируемых структур. Однако понимание структурных принципов в той мере, которая необходима для проведения таких различий, пока еще не достигнуто. Вместо случайным образом генерированных свернутых цепей можно использовать структурно изученные белки. Однако число известных типов свертывания цепей для этого недостаточно велико.
Матрица сравнения Са -расстояний может выявить совпадение цепей. Структуры отдаленно родственных белков обычно сильно отличаются друг от друга зa счет больших вставок и делений, как это показано на рис. 9.4 для структур химотрипсина и протеазы В. При сравнении свертывания цепей, как и при сравнении аминокислотных последовательностей, требуется совпадение соответствующих остатков, хотя на среднеквадратичном Са-расстоянии нарушения совпадения сказываются не так резко, как на сумме М. Такое совмещение можно производить по той же схеме, что и при сравнении последовательностей, рис. 9.5, б; М(р, q) нужно заменить среднеквадратичными Са -расстояниями между сегментами цепи данной длины, центрированными относительно р и q [802]. После установления совпадений можно определить общее среднеквадратичное расстояние между соответствующими остатками. Однако его опять-таки нельзя перевести в стандартную значимость подобия двух рассматриваемых белков, поскольку кумулятивные вероятности для среднеквадратичных Са-расстояний пока не известны.
Матрица сравнения среднеквадратичных Са-расстояний также может быть использована для выявлений структурного подобия между частями белков. Такое подобие обнаружено, например, между фрагментами лизоцима бактериофага Т4 и лизоцима белка куриного яйца [802]. Анализируемые длины сегментов составляли 40 и 80 остатков.
Сравнения укладки цепи должны учитывать вставки и делеции.
При расчете среднеквадратичных Са-расстояний между фрагментами цепи, которые сопоставляют с помощью матрицы сравнения, вставками и делениями полностью пренебрегают. Такое допущение, однако, слишком произвольно, поскольку информация о вставках и делениях необходима для определения показателя подобия. Действительно, если в сравниваемых белках их оказывается слишком много, соответствующие белки считают не проявляющими подобия. При попарном сравнении свертываний цепи вставки и делеции можно учесть с соответствующими весами путем нахождения среднего Са-расстояния по минимальной площади, заключенной между сравниваемыми цепями, как показано на рис. 9.6. Этот метод не требует совпадения. Однако минимизация площади, заключенной между двумя цепями, в вычислительном отношении более сложна, чем минимизация расстояний между двумя одинаковыми по величине наборами атомов, которые составляют базис при определении среднеквадратичного Са-расстояния.
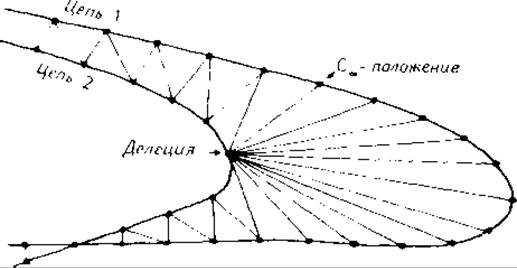
Рис. 9.6. Расчет среднего Са-расстояния между двумя цепями, учитывающий систематические вставки и делеции [387].
Среднее Са-расстояние определяется как минимальная площадь, заключенная между двумя цепями (соответствующая площади эластичной пленки, натянутой между двумя проволоками) и отнесенная к средней длине цепи. Сумма площадей треугольников между Са-атомами, которые показаны на рисунке, является хорошей аппроксимацией этой минимальной площади.