БИОХИМИЯ - Л. Страйер - 1984
ТОМ 1
ЧАСТЬ I. КОНФОРМАЦИЯ И ДИНАМИКА
ГЛАВА 2. ОСНОВНЫЕ ПРЕДСТАВЛЕНИЯ О СТРУКТУРЕ И ФУНКЦИИ БЕЛКОВ
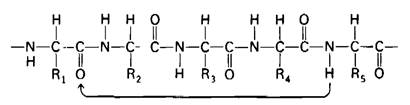
2.8. Конформация полипептидных цепей
Поразительная особенность белков состоит в том, что каждый из них имеет четко определенную трехмерную структуру. Как показано ниже, будучи развернутыми или уложенными случайным образом, полипептидные цепи лишены биологической активности. Функциональные свойства белков определяются их конформацией, т. е. пространственным расположением атомов. Важную роль при этом играет последовательность аминокислот, так как в конечном счете именно она определяет конформацию белка.
В конце 30-х годов Л. Полинг (L. Pauling) и Р. Кори (R. Corey) начали проводить рентген оструктурные исследования аминокислот и петидов. Они определяли стандартные длины и углы связей, с тем чтобы исходя из этих данных предсказать конформацию белков. Был обнаружен важный факт: пептидная единица обладает жесткой планарной (плоской) структурой. Водород в замещенной аминогруппе почти всегда занимает трансположение по отношению к кислороду карбонильной группы (рис. 2.33). Связь между атомом углерода карбонильной группы и атомом азота пептидной единицы имеет частично характер двойной связи, и, следовательно, вращение вокруг этой (рис. 2.34) связи должно быть заторможено. Длина связи составляет 1,32 А-среднее значение между длинами одинарной связи С — N (1,49 А) и двойной связи C=N (1,27 А).
Рис. 2.33. Пептидная группа имеет жесткую планарную структуру. Показаны длины связей (в А)
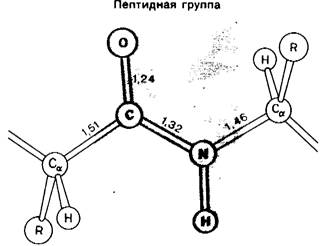
Рис. 2.34. Планарность пептидной группы обусловлена тем, что связь азот-углерод носит частично характер двойной связи

Ангстрем (А)-единица длины, равная 10-10 м. 1 А= 10-10 м = 10-8 см = 10-4 мкм = = 10-1 нм. Названо в честь ученого-спектроскописта А. Ангстрема (1814-1874).
В отличие от рассмотренного случая связь между α-углеродным атомом и углеродным атомом карбонильной группы истинно одинарная. Следовательно, по обеим сторонам жесткой пептидной единицы вокруг этих связей имеется высокая степень свободы вращения (рис. 2.35). Вращения относительно этих связей описываются углами ![]() и ф.(рис. 2.36).
и ф.(рис. 2.36).
Рис. 2.35. В области связей между пептидными группами и α-углеродными атомами имеется довольно высокая степень свободы вращения
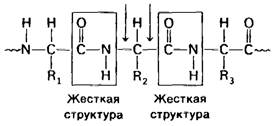
Рис. 2.36. Определение углов ![]() и ф:
и ф: ![]() характеризует вращение относительно одинарной связи Сα —С; ф характеризует вращение относительно одинарной связи Сα—N. (Levintha! СMolecular model building by computer, Scientific American, Inc., 1966.)
характеризует вращение относительно одинарной связи Сα —С; ф характеризует вращение относительно одинарной связи Сα—N. (Levintha! СMolecular model building by computer, Scientific American, Inc., 1966.)
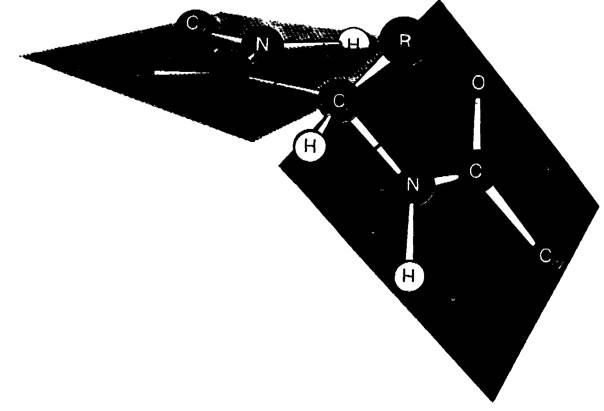
Для полного описания конформации основной цепи полипептида необходимо знать ![]() для каждого аминокислотного остатка.
для каждого аминокислотного остатка.
2.9. Периодичные структуры: альфа-спираль, бета-складчатый слой, спираль коллагена
Может ли полипептидная цепь быть уложена в структуру, состоящую из регулярно повторяющихся участков? Чтобы ответить на этот вопрос, Полинг и Кори сравнили ряд потенциально возможных конформаций полипептидов, построив их точные молекулярные модели. При этом строго соблюдались экспериментально установленные для аминокислот и небольших пептидов величины углов связей и межатомных расстояний. В 1951 г. они предложили две периодические полипептидные структуры, названные соответственно α-спираль и β-складчатый слой.
α-Спираль имеет вид стержня. Туго закрученная основная цепь полипептида создает внутреннюю часть стержня, а боковые цепи направлены наружу от основной цепи, располагаясь по спирали (рис. 2.37 и 2.38). α-Спираль стабилизирована водородными связями между NН- и СО-группами основной цепи. СО-группа каждой аминокислоты соединяется водородной связью с NН-группой аминокислоты, расположенной в линейной последовательности на 4 остатка впереди (рис. 2.39). Таким образом, все СО- и NН-группы основной цепи связаны между собой водородными связями. В проекции на ось спирали один остаток отстоит от другого на 1,5 А, а угол между ними составляет 100, т.е. на полный виток спирали приходится 3,6 аминокислотных остатка. Таким образом, аминокислоты, отстоящие друг от друга на 3-4 остатка в линейной последовательности, в структуре α-спирали пространственно расположены очень близко друг к другу. Напротив, аминокислоты, разделенные в линейной последовательности двумя остатками, пространственно располагаются на противоположных сторонах спирали и поэтому взаимодействие между ними маловероятно. Шаг α-спирали составляет 5,4 А2 расстояние между остатками (по оси)-1,5 А и число остатков на один оборот-3,6. Спираль может закручиваться по часовой стрелке (правая спираль) или против часовой стрелки (левая спираль); все исследованные а-спирали белков относятся к правому типу.
Рис. 2.37. Модель правозакрученной α-спирали. А. На спирали показаны только а-углеродные атомы. Б. Показаны только атомы азота (N), образующие скелет молекулы, атомы а- углерода (Са) и карбонильного углерода (С). В. Полное изображение спирали. Водородные связи (на рис. В обозначены красными точками) между NH- и СО-группами стабилизируют спираль
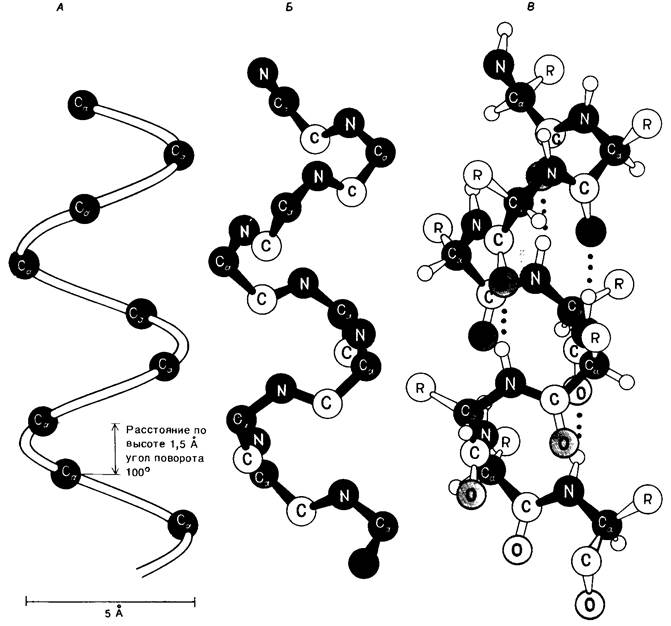
Рис. 2.38. α-Спираль в поперечном разрезе. Обратите внимание, что боковые цепи (показаны зеленым) находятся снаружи спирали. Вандерваальсовы радиусы атомов на самом деле больше, чем изображено на рисунке, вследствие этого внутри спирали почти нет свободного пространства
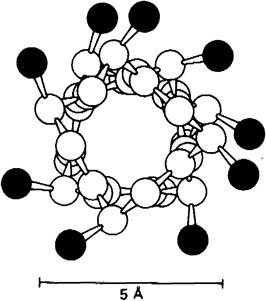
Рис. 2.39. В α-спирали NР-группа n-го остатка связана водородной связью с СО-группой остатка (n- 4)
Содержание α-спиралей в белках, изученных к настоящему времени, крайне вариабельно. В некоторых белках, например, миоглобине и гемоглобине, α-спираль лежит в основе структуры. Другие белки, например, пищеварительный фермент химотрипсин, практически лишены α-спиральной структуры. Одинарная α-спираль, о которой речь шла выше, как правило, довольно коротка, обычно менее 40 А в длину. Варианты α-спиралей используются при образовании длинных тяжей, достигающих 1000 А и более в длину. Две или более α-спирали могут закручиваться одна вокруг другой, как тяжи в канате. Такая структура-α-спирализованная суперспираль- обнаружена во многих белках; в кератине волос, миозине и тропомиозине мышц, эпидермисе кожи и фибрине, в сгустках крови. Спирализованные «канаты» этих белков выполняют механическую роль, образуя плотные пучки волокон.
«Если принять, что фибриллярные белки эпидермиса, белки кератинизированных тканей, основной белок мышц миозин, а теперь и фибриноген крови-все имеют в основе одну и ту же особую форму молекулярной структуры и потому, вероятно, представляют собой адаптационные варианты одного исходного принципа, то здесь мы, видимо, столкнулись с одним из великих фактов эволюции биологических молекул».
К. Bailey, W.T. Astbury, К. М. Rudall, Nature, 1943
Структура α-спирали была предсказана Полингом и Кори за 6 лет до того, как ее удалось экспериментально выявить методом рентгеноструктурного анализа миоглобина. Открытие структуры α -спирали представляет собой важную веху в развитии молекулярной биологии, поскольку это открытие доказало, что можно предсказать конформацию полипептидной цепи, если точно известны свойства ее компонентов.
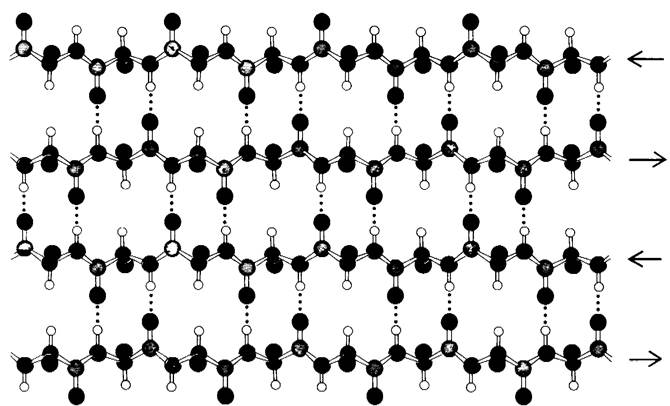
В тот же год Полинг и Кори открыли другой вариант периодической структуры, который они назвали β-складчатым слоем β потому, что это была вторая-после α-спирали - открытая ими структура), β-складчатый слой существенно отличается от α- спирали тем, что он имеет плоскую, а не стержневидную форму. Полипептидные цепи в β-складках почти полностью вытянуты (рис. 2.40), а не туго скручены, как в α-спирали. Расстояние по оси между двумя прилежащими аминокислотами составляет 3,5, а не 1,5 А, как в α-спирали. Другая особенность β-складчатой структуры состоит в том, что она стабилизирована водородными связями между NH- и СО-группами разных тяжей полипептидных цепей, тогда как в а-спирали водородные связи образуются между этими группами в пределах одной и той же полипептидной цепи. Прилежащие цепи в складчатом β-слое могут идти в одном и том же направлении (параллельный β-слой) или в противоположных направлениях (антипараллельный β-слой). Например, фиброин шелка состоит почти целиком из «штабелей» антипараллельных β-складчатых слоев (рис. 2.41). Аналогичные области β-склалчатых слоев встречаются во многих других белках. Особенно широко распространены структурные единицы, состоящие из 2-5 параллельных или антипараллельных β-складок.
Рис. 2.40. Конформация дипептидной единицы в складчатом β-слое. Полипептидная цепь при этом практически полностью вытянута
Рис. 2.41. Антипараллельный β-складчатый слой, Прилежащие тяжи полипептидной цепи идут в противоположных направлениях. Структура стабилизирована водородными связями между NН- и СО-группами прилежащих тяжей. Боковые цепи (показаны зеленым) лежат выше и ниже плоскости слоя
Третий тип периодической структуры - коллагеновая спираль-будет подробно рассматриваться в гл. 9. Эта специализированная структура обеспечивает высокую упругость коллагена-основного компонента кожи, костей и сухожилий.