БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 13. ЦИКЛ ТРИКАБОНОВЫХ КИСЛОТ
13.17. Регуляция пируват-дегидрогеназного комплекса
Образование ацетил-СоА из пирувата-это ключевой необратимый этап метаболизма, потому что животные неспособны к превращению ацетил-СоА в глюкозу. Окислительное декарбоксилирование пирувата в ацетил-СоА создает потенциальную возможность для дальнейших превращений атомов глюкозы по двум путям: I) окисление до СO2 через цикл трикарбоновых кислот с одновременным генерированием энергии или 2) включение в липиды (разд. 17.21). Следовательно, есть все основания думать, что активность пируват-дегидрогеназного комплекса должна строго регулироваться. И действительно, этот ферментный комплекс регулируется тремя путями.
1. Ингибирование продуктами реакции. Ацетил-СоА и NАDН, продукты окисления пирувата, ингибируют ферментный комплекс. Ацетил-СоА подавляет активность трансацетилазного компонента, а N АDН - активность дигидролипоил-дегидрогеназного компонента. Эти ингибиторные эффекты обращаются при действии соответственно СоА и NАD+.
2. Регуляция нуклеотидами по принципу обратной связи. Активность ферментного комплекса регулируется энергетическим зарядом (разд. 11.13). Пируват-дегидроге- назный компонент специфически ингибируется GТР и активируется АМР. Следовательно, активность комплекса снижается, когда клетка богата легкодоступной энергией.
3. Регуляция путем ковалентной модификации. Комплекс теряет ферментативную активность, когда специфический остаток серина пируват-дегидрогеназного компонента фосфорилируется АТР. Фосфорилирование усиливается при высоких соотношениях ATP/ADP, ацетил-СоА/СоА, NADH/NAD+ и ингибируется пируватом. Ферментный комплекс вновь активируется, если фосфорильная группа гидролизуется специфической фосфатазой. Дефосфорилирование усиливается при высоком содержании пирувата. Ковалентная модификация представляет собою важный механизм регуляции ферментативной активности. Мы вновь встретимся с регуляторной ролью реакций фосфорилирования и дефосфорилирования, когда будем рассматривать синтез и распад гликогена.
13.18. Регуляция цикла трикарбоновых кислот
Скорость функционирования цикла трикарбоновых кислот точно пригнана к потребности клеток в АТР. Важной регуляторной реакцией цикла является синтез цитрата из оксалоацетата и ацетил-СоА. АТР-аллостерический ингибитор цитрат-синтазы. Его действие заключается в повышении Км для ацетил-СоА. Таким образом, с увеличением содержания АТР снижается насыщение фермента ацетил-СоА и в результате уменьшается образование цитрата.
Вторая регуляторная реакция-это реакция, катализируемая изоцитрат-дегидрогеназой. Фермент аллостерически стимулируется ADP, который повышает его сродство к субстратам. Между связыванием изоцитрата, NAD+, Mg2+ и ADP существует взаимная кооперативность. В отличие от этого NADH ингибирует изоцитратдегидрогеназу путем прямого вытеснения NAD+.
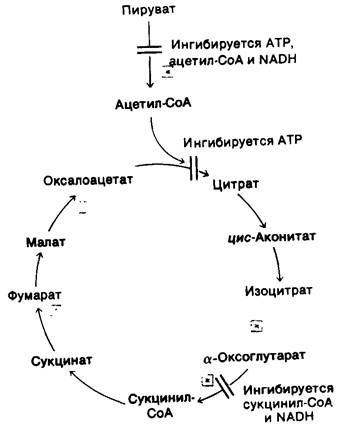
Третьей регуляторной реакцией цикла трикарбоновых кислот является реакция, катализируемая α-оксоглутарат—дегидрогеназой. Регуляция на этом этапе в некоторых отношениях подобна регуляции на уровне пируват-дегидрогеназного комплекса, как и можно было ожидать, исходя из их структурной гомологии. α-Оксоглутарат - дегидрогеназа ингибируется сукцинил-СоА и NADH, т. е. продуктами катализируемой ею реакции. Она ингибируется также высоким энергетическим зарядом. Коротко говоря, поступление двухуглеродных фрагментов в цикл трикарбоновых кислот и скорость цикла снижаются при высоком содержании АТР в клетке. Эта регуляция достигается участием ряда комплементарных механизмов на различных стадиях цикла (рис. 13.16).
Рис. 13.16. Регуляция цикла трикарбоновых кислот и окислительного декарбоксилирования пирувата: звездочкой в квадрате указаны стадии, на которых требуется акцептор электронов (NAD+ или FAD), регенерируемый в дыхательной цепи
13.19. Открытие цикла Кребсом
«Меня часто спрашивали, как возникло и развивалось исследование цикла трикарбоновых кислот. Была ли эта концепция результатом внезапного вдохновения и предвидения?» «Ничего подобного, отвечает Ганс Кребс (Hans Krebs), это был очень медленный эволюционный процесс, развивавшийся в течение пяти лет начиная с 1932 г. (когда я включился в эту работу)...» Кребс вначале изучал скорость окисления различных соединений, используя срезы почек и печени. Он выбирал вещества, представлявшие собою возможные промежуточные продукты при окислении пищевых веществ. Кребс предполагал, что такие вещества будут быстро окисляться, а, следовательно, их легко будет идентифицировать. Были получены важные данные: цитрат, сукцинат, фу- марат и ацетат очень быстро окислялись в различных тканях.
Существенный вклад в изучение этой проблемы внес Альберт Сент-Дьёрдьи (Albert Szent-Györgyi) в 1935 г. Он исследовал окисление различных веществ, используя суспензии измельченной грудной мышцы голубя. Эта очень активная летательная мышца обладает исключительно высокой скоростью окислительных процессов, что ускоряло ход эксперимента. Сент-Дьёрдьи обнаружил, что добавление некоторых С4-дикарбоновых кислот повышало потребление О2 в гораздо большей степени, чем это требовалось для их прямого окисления. Иными словами, они каталитически (а не стехиометрически) увеличивали потребление О2. Такая каталитическая стимуляция дыхания наблюдалась в присутствии сукцината, фумарата и малата.
Следующим шагом вперед явилась расшифровка биологического пути окисления цитрата Карлом Мартиусом и Францем Кноопом (Carl Martius, Franz Knoop) в 1937 г. Они показали, что цитрат изомери- зуется в изоцитрат через цис-аконитат и что изоцитрат подвергается окислительному декарбоксилированию в α-оксоглутарат. Возможность окисления а-оксоглутарата в сукцинат тогда уже была известна, и, таким образом, это открытие позволило установить путь от цитрата до сукцината. Оно подоспело в нужный момент, так как теперь Кребс смог объяснить свое недавнее наблюдение, что цитрат каталитически усиливает дыхание измельченной грудной мышцы голубя.

Дополнительная важная информация была получена в результате использования малоната, специфического ингибитора сукцинат-дегидрогеназы. Малонат является конкурентным ингибитором этого фермента, поскольку он структурно очень близок сукцинату. К этому времени было известно, что малонат служит дыхательным ядом. Кребс пришел к выводу, что сукцинат-дегидрогеназа может, таким образом, играть ключевую роль в дыхании. В пользу этого предположения говорил тот факт, что при добавлении цитрата к мышце, отравленной малонатом, в ней накапливается сукцинат. Более того, накопление сукцината в такой мышце происходило также при добавлении фумарата. Первый из этих экспериментов указывает на физиологическую значимость пути от цитрата до сукцината. Второй эксперимент раскрывает существование пути от фумарата до сукцината, отличающегося от реакции, катализируемой сукцинат-дегидрогеназой.
Кребс установил далее, что цитрат быстро образуется в мышечной суспензии при добавлении оксалоацетата. Открытие синтеза цитрата из оксалоацетата позволило Кребсу построить полную схему процесса. Постулированный им цикл трикарбоновых кислот сразу создал ясную картину окисления углеводов. В этой картине нашли свое
точное место многие экспериментальные факты-такие, например, как каталитическое усиление дыхания сукцинатом и другими промежуточными продуктами. Примечательно, что цикл трикарбоновых кислот-не единственный и не первый метаболический цикл, раскрытый Кребсом. Шестью годами ранее он показал, что мочевина превращается по циклическому метаболическому пути, названному орнитиновым циклом (гл. 18). Таким образом, концепция циклического метаболического пути уже была полностью осознана Кребсом, когда он анализировал данные и определял ход экспериментов, которые привели к предложенному им циклу трикарбоновых кислот.