БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 14. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ
NADH и FADH2, образовавшиеся при гликолизе, окислении жирных кислот и в цикле трикарбоновых кислот, представляют собою богатые энергией молекулы, поскольку каждая из них содержит пару электронов с высоким потенциалом переноса. При переносе этих электронов на молекулярный кислород высвобождается большое количество энергии. Высвободившаяся энергия может быть использована для генерирования АТР. Окислительное фосфорилирование - это процесс образования АТР, сопряженного с транспортом электронов по цепи переносчиков от NADH или FADH2 к O2. У аэробных организмов этот процесс служит главным источником АТР. Так, окислительное фосфорилирование обеспечивает генерирование 32 молекул АТР из 36, образующихся в процессе полного окисления глю
козы до СO2 и Н2O. Наиболее характерные черты этого процесса сводятся к следующему.
1. Окислительное фосфорилирование осуществляется дыхательными ансамблями, локализованными во внутренней мембране митохондрий. Цикл трикарбоновых кислот и путь окисления жирных кислот, поставляющие большую часть NADH и FADH2, осуществляются в прилежащем митохондриальном матриксе.
2. Окисление NADH дает 3 АТР, тогда как окисление FADH2 дает 2 АТР. Окисление и фосфорилирование-процессы сопряженные.
Дыхание - процесс генерирования АТР, в котором роль конечного акцептора электронов выполняет неорганическое соединение (такое, как O2). Донором электронов может служить как органическое, так и неорганическое соединение.
Рис. 14.1. Электронная микрофотография митохондрии
3. Дыхательные ансамбли включают в себя многочисленные переносчики электронов, в частности цитохромы. Многоступенчатый транспорт электронов от NADH или FАDН2 к O2 по цепи этих переносчиков приводит к выбросу протонов из митохондриального матрикса и генерированию мембранного потенциала (протонодвижущей силы). Протоны перекачиваются тремя видами электронпереносящих комплексов. В процессе обратного тока протонов в митохондриальный матрикс при посредстве
ферментного комплекса происходит синтез ATP. Таким образом, сопряжение окисления и фосфорилирования обеспечивается протонным градиентом через внутреннюю митохондриальную мембрану.
14.1. Окислительное фосфорилирование происходит в митохондриях
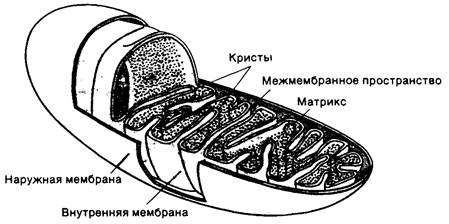
Митохондрии представляют собою органеллы овальной формы длиной обычно около 2 мкм и диаметром 0,5 мкм. Метод выделения митохондрий был разработан в конце 40-х годов. Юджин Кеннеди и Элберт Ленинджер (Eugene Kennedy, Albert Lehninger) открыли тогда присутствие в митохондриях дыхательных ансамблей, ферментов цикла трикарбоновых кислот и ферментов окисления жирных кислот. Электронно-микроскопические исследования, выполненные Георгом Паладом и Фритьофом Сьёстрандом (George Palade, Fritjof Sjöstrand), показали, что митохондрии имеют две мембранные системы: наружную мембрану и имеющую большую поверхность внутреннюю мембрану. Последняя образует многочисленные глубокие складки (гребневидные выросты), называемые кристами. Таким образом, в митохондриях имеются два компартмента: межмембранное пространство (между наружной и внутренней мембранами) и матрикс, ограниченный внутренней мембраной (рис. 14.2). Дыхательные ансамбли составляют неотьемлемую часть внутренней митохондриальной мембраны, тогда как большая часть реакций цикла трикарбоновых кислот и окисления жирных кислот протекает в матриксе.
Наружная мембрана вполне проницаема для большинства мелких молекул и ионов. В противоположность этому внутренняя мембрана проницаема почти для всех ионов и для большинства незаряженных молекул. Существуют специальные белковые переносчики, транспортирующие через внутреннюю митохондриальную мембрану такие молекулы, как АТР и жирные кислоты с длинной цепью.
Рис. 14.2. Схематическое изображение митохондрий