БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 16. ГЛИКОГЕН И ОБМЕН ДИСАХАРИДОВ
16.2. Для распада гликогена необходим также фермент, разрывающий связи в местах ветвления
Под действием одной только фосфорилазы гликоген распадается в ограниченной степени. α-1,6-гликозидные связи в точках ветвления нечувствительны к расщеплению фосфорилазой, поскольку ее действие на α-1,4-связи приостанавливается по достижении концевого остатка, отстоящего от точки ветвления на четыре остатка. Действие фосфорилазы на две внешние ветви гликогеновой частицы показано на рис. 16.4. Пять α-1,4-гликозидных связей на одной ветви и три на другой расщепляются фосфорилазой. На этой стадии процесс приостанавливается, поскольку концевые остатки а и г удалены от точки ветвления з на четыре остатка. На этом этапе требуется подключение новой ферментативной активности. Трансфераза переносит блок из трех гликозильных остатков с одной внешней ветви на другую. α-1,4-гликозидная связь между в и Z разрывается и образуется новая α-1,4-связь между в и г. В результате такого переноса остаток Z обнажается и становится доступным действию третьего деградирующего фермента, α-1,6- глюкозидазы, известной также как дебранчинг-фермент. Этот фермент гидролизует α-1,6-гликозидную связь между остатками Z и з.
Фосфоролиз -расщепление связи ортофосфатом (в отличие от гидролиза, означающего расщепление водой).
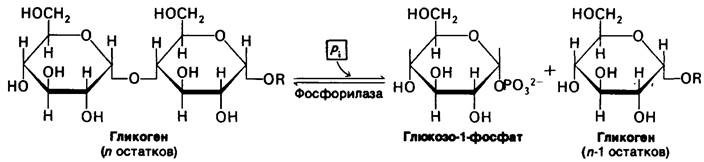
Рис. 16.4. Стадии расщепления гликогена
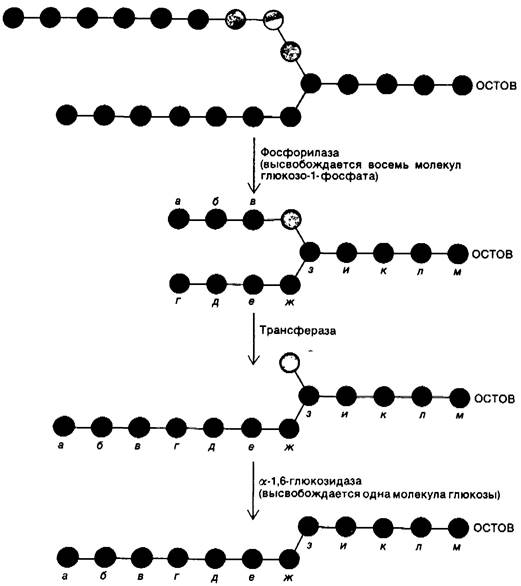
Таким образом, трансфераза и фермент, разрывающий связи в местах ветвления (α-1,6-глюкозидаза), превращают разветвленную структуру в линейную, что прокладывает путь для дальнейшего расщепления молекулы фосфорилазой. В результате отщепления Z все остатки от а до м становятся чувствительными к фосфорилазе. Интересно отметить, что одна полипептидная цепь в 160 к Да содержит и трансферазный, и α-1,6-глюкозидазный активные центры.
16.3. Фосфоглюкомутаза превращает глюкозо-1-фосфат в глюкозо-6-фосфат
Глюкозо-1-фосфат, образовавшийся при фосфоролитическом расщеплении гликогена, превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Равновесная смесь содержит 95% глюкозо-6-фосфата. В каталитическом центре активной молекулы фермента присутствует фосфорилированный остаток серина. Во время катализа эта фосфорильная группа, вероятно, переносится на гидроксильную группу при С-6 глюкозо-1-фосфата с образованием глюкозо-1,6-бисфосфата. Далее фосфорильная группа указанного промежуточного продукта переносится на остаток серина в активном центре. В результате происходит образование глюкозо-6-фосфата и регенерирование фосфорилированного фермента.
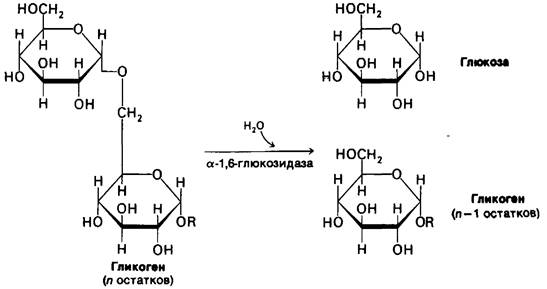
Фосфорильная группа мутазы медленно теряется в процессе гидролиза. Она восстанавливается благодаря переносу фосфорильной группы от глюкозо-1,6-бисфосфата, который образуется из глюкозо-1-фосфата и АТР в реакции, катализируемой фосфоглюкокиназой. Эти процессы подобны реакциям, катализируемым гликолитическим ферментом фосфоглицеромутазой (разд. 12.7). Роль 2,3-бисфосфоглицерата (2,3-БФГ) во взаимопревращении 2-фосфоглицерата и 3-фосфоглицерата подобна таковой глкжозо-1,6-бисфосфата во взаимопревращениях фосфоглюкоз. Кроме того, в обеих реакциях промежуточным продуктом служит фосфорилированный фермент.
16.4. Печень содержит глюкозо-6-фосфатазу, гидролитический фермент, отсутствующий в мышцах
Главная функция печени, это поддерживать содержание глюкозы в крови на относительно постоянном уровне. Печень высвобождает глюкозу в кровь в период мышечной активности и в интервалах между приемами пищи. Высвобождаемая глюкоза поглощается преимущественно мозгом и скелетными мышцами. Фосфорили- рованная глюкоза в противоположность неэтерифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глнжозо-6-фосфатазу, который и обеспечивает возможность выхода глюкозы из этого органа. Этот фермент играет важную роль в глюконеогенезе (разд. 15.14).
Глюкозо-6-фосфат + Н2O → Глюкоза + Pi.
Глюкозо-6-фосфатаза имеется также в почках и кишечнике, но ее нет в мышцах и мозгу. Следовательно, глюкозо-6-фосфат задерживается мышцами и мозгом, которые нуждаются в больших количествах этого топлива для генерирования АТР. В то же время для печени глюкоза не является главным топливом. Печень запасает и высвобождает глюкозу прежде всего для удовлетворения потребности в ней других тканей (разд. 23.5).