БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 16. ГЛИКОГЕН И ОБМЕН ДИСАХАРИДОВ
16.5. Синтез и расщепление гликогена происходят различными путями
Реакция, катализируемая гликоген-фосфорилазой, легко обратима, потому что ∆G0 для элонгации гликогена с участием глюкозо-1-фосфата составляет —0,5 ккал/моль. Действительно, Кори удалось синтезировать гликоген из глюкозо-1-фосфата с использованием фосфорилазы и ветвящего фермента. Однако ряд последующих экспериментальных исследований показал, что in vivo гликоген синтезируется иным путем. Во-первых, реакция, катализируемая фосфорилазой, находится в состоянии равновесия, когда отношение [Рi]/[Глюкозо-1-фосфат] равно 3,6 при нейтральном значении pH, тогда как в клетках это отношение обычно превышает 100. Следовательно, in vivo фосфорилазная реакция должна идти в направлении распада гликогена. Во-вторых, гормоны, вызывающие повышение активности фосфорилазы, обычно усиливают распад гликогена. В-третьих, больные с полным отсутствием в мышцах фосфорилазы [болезнь Мак-Ардля (McArdle)] (разд. 16.19) способны к синтезу мышечного гликогена.
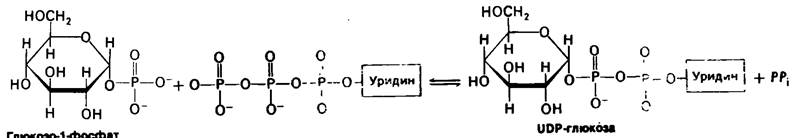
В 1957 г. Луи Лелуа (Luis Leloir) и его сотрудники показали, что синтез гликогена происходит другим путем. Донором гликозильной группы служит уридиндифосфат- глюкоза (UDP-глюкоза), а не глюкозо-1- фосфат. Реакция синтеза гликогена, это не обращенная реакция его расщепления:
Синтез: Гликогенn + UDP-глюкоза → Гликоген n+1 + UDP.
Распад: Гликоген n+1 + Pi → Гликоген n + Глюкозо-1-фосфат.
Мы знаем теперь, что пути биосинтеза и расщепления в биологических системах почти всегда различны. Обмен гликогена— первый пример этого важного принципа. Существование раздельных путей синтеза и распада обеспечивает значительно большую гибкость процессов, как в энергетическом, так и в регуляторном плане. Клетка не отдается больше только «на милость» закона действия масс. Гликоген может синтезироваться вопреки высокому соотношению [Ортофосфат]/[Глюкозо-1-фосфат].
16.6. UDP-глюкоза-активированная форма глюкозы
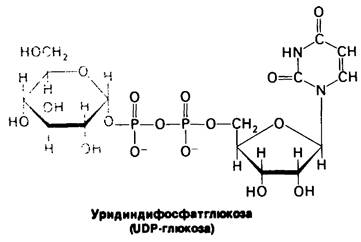
UDP-глюкоза, донор глюкозы в биосинтезе гликогена, представляет собою активированную форму глюкозы, аналогично тому как АТР и ацетил-СоА являются активированными формами ортофосфата и ацетата соответственно. Атом углерода С-1 гликозильного компонента UDP-глюкозы активирован, потому что его гидроксильная группа образует эфирную связь с дифосфатным компонентом UDP.
UDP-глюкоза синтезируется из глюкозо-1-фосфата и уридинтрифосфата (UTP) в ходе реакции, катализируемой UDP-глюкозо-пирофосфорилазой. Пирофосфат, освобождающийся при этой реакции, происходит из двух обращенных наружу фосфорильных остатков UTP.
Реакция легко обратима, но пирофосфат быстро гидролизуется in vivo в ортофосфат неорганической пирофосфатазой. Необратимый по своей природе гидролиз пирофосфата запускает синтез UDP-глюкозы.
Глюкозо-1-фосфат + UTP ⇄ UDP-глюкоза + PP¡,
PPi + Н2O → 2Pi
Глюкозо-1-фосфат + UTP + Н2O → UDP-глюкоза + 2Pi
Синтез UDP-глюкозы может служить примером часто повторяющейся в биохимии темы; многие реакции биосинтеза запускаются гидролизом пирофосфата. Широкое значение имеет другой аспект этой реакции. Нуклеозиддифосфатные сахара служат донорами гликозильной группы в биосинтезе многих дисахаридов и полисахаридов.
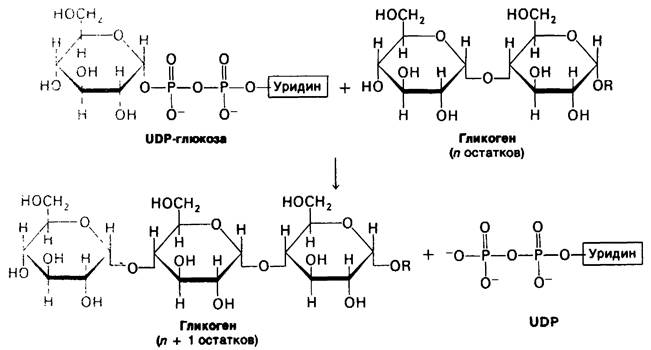
16.7. Гликоген-синтаза катализирует перенос глюкозы от UDP-глюкозы к растущей цепи
Новые глюкозильные фрагменты присоединяются к невосстанавливающим концевым остаткам гликогена. Активированный глюкозильный компонент UDP- глюкозы переносится на гидроксильную группу C-4-конца гликогена, образуя α-1,4- гликозидную связь. В происходящей реакции элонгации UDP замещается этой концевой гидроксильной группой растущей молекулы гликогена. Реакция катализируется гликоген-синтазой, осуществляющей присоединение глюкозильных остатков только при условии, что полисахаридная цепь уже содержит более четырех остатков. Таким образом, для синтеза гликогена требуется наличие затравки, образуемой другой синтетазой.