БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 17. ОБМЕН ЖИРНЫХ КИСЛОТ
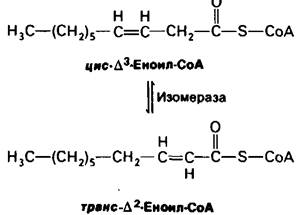
17.10. Необходимость изомеразы и эпимеразы для окисления ненасыщенных жирных кислот
Обратимся теперь к окислению ненасыщенных жирных кислот. Многие из происходящих при этом реакций идентичны таковым для насыщенных жирных кислот. Действительно, для расщепления большого числа ненасыщенных жирных кислот требуются только два добавочных фермента-изомераза и эпимераза.
Рассмотрим окисление пальмитолеата. Эта ненасыщенная жирная С16-кислота, имеющая одну двойную связь между С-9 и С-10, активируется и транспортируется через внутреннюю митохондриальную мембрану таким же образом, как пальмитат. Пальмитолеил-СоА претерпевает далее три цикла расщепления, катализируемые теми же ферментами, что и окисление насыщенных жирных кислот. Однако цис-∆3-еноил-СоА, образуемый в третьем цикле, не является субстратом для ацил- СоА -дегидрогеназы. Наличие двойной связи между С-3 и С-4 предотвращает образование другой двойной связи, между С-2 и С-3. Это препятствие преодолевается с помощью новой реакции, которая сдвигает положение и конфигурацию цис-∆3-двойной связи. Под действием изомеразы эта двойная связь превращается в транс-А2-двойную связь. Последующие реакции подобны реакциям пути окисления насыщенных жирных кислот, когда транс-∆2-еноил-СоА является обычным субстратом.
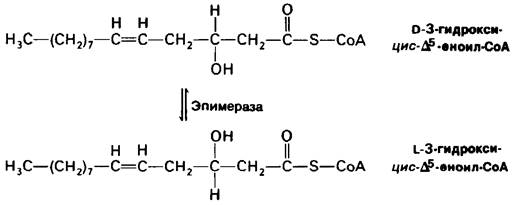
Для окисления полиеновых жирных кислот необходим и второй добавочный фермент. Активированная ненасыщенная жирная С18-кислота с двойными цис-∆16- и цис-∆9-связями претерпевает два цикла деградации по пути окисления насыщенных жирных кислот. Затем цис-∆2. Д5-еноил-СоА гидратируется еноил-СоА - гидратазой, тем же ферментом, который гидратирует двойную транс-∆2-связь при окислении насыщенных жирных кислот. Однако гидратирование двойной цис-∆2-связи дает D-изомер 3-гидроксиацил-СоА, который не может служить субстратом для L-З-гидроксиацил- СоА-дегидрогеназы. Это затруднение снимается эпимеразой, обращающей конфигурацию гидроксильной группы при С-3.
17.11. Жирные кислоты с нечетным числом углеродных атомов образуют при завершающем тиолизе пропионил-СоА
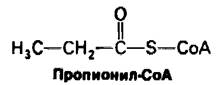
Жирных кислот с нечетным числом углеродных атомов насчитывается немного. Они окисляются таким же образом, как и жирные кислоты с четным числом атомов, с той лишь разницей, что на последнем этапе расщепления образуются одна молекула пропионил-СоА и одна молекула ацетил- СоА, а не две молекулы ацетил-СоА. Активированный трехуглеродный фрагмент пропионил-СоА включается в цикл трикарбоновых кислот после превращения в сукцинил-СоА. Путь от пропионил-СоА к сукцинил-СоА рассматривается в следующей главе (разд. 18.11), поскольку пропионил- СоА образуется также при окислении некоторых аминокислот.
17.12. Образование кетоновых тел из ацетилкофермента А в условиях, когда расщепление жиров преобладает
Ацетил-СоА, образовавшийся при окислении жирных кислот, включается в цикл трикарбоновых кислот в условиях, когда расщепление жиров и углеводов соответствующим образом сбалансировано. Теперь уже ясна молекулярная основа изречения, что жиры сгорают в пламени углеродов. Включение ацетил-СоА в цикл зависит от доступности оксалоацетата для образования цитрата. Однако, если расщепление жиров преобладает, судьба ацетил-СоА изменяется. Объясняется это тем, что в отсутствие углеводов или при нарушении их использования концентрация оксалоацетата снижается. В случае голодания или диабета оксалоацетат расходуется на образование глюкозы и поэтому не может конденсироваться с ацетил-СоА. В таких условиях путь метаболизма ацетил-СоА отклоняется в сторону образования ацетоацетата и D-З-гидроксибути- рата, Ацетоацетат, D-З-гидроксибутират и ацетон часто называют кетоновыми телами.
Ацетоацетат образуется из ацетил-СоА в три стадии. Две молекулы ацетил-СоА конденсируются с образованием ацетоацетил-СоА. Эта реакция, катализируемая тиолазой, представляет собою обращение тиолизной стадии в окислении жирных кислот. Ацетоацетил-СоА реагирует далее с ацетил-СоА и Н2O, образуя З-гидрокси-З-метилглутарил-СоА и СоА. Неблагоприятное равновесие в образовании ацетоацетил-СоА компенсируется благоприятным равновесием этой последней реакции, обусловленным гидролизом тиоэфирной связи. З-гидрокси-З-метилглутарил- СоА затем расщепляется на ацетил-СоА и ацетоацетат. Суммарная реакция этих пре-
вращений имеет следующий вид:
2 Ацетил-СоА + Н2O→ Ацетоацетат + 2СоА + Н+.
При восстановлении ацетоацетата в митохондриальном матриксе образуется 3-гидроксибутират. Отношение гидроксибутирата к ацетоацетату зависит от отношения [NADH]/[NAD+ ] в митохондриях. Ацетоацетат претерпевает также медленное спонтанное декарбоксилирование в ацетон. У людей с высоким содержанием ацетоацетата в крови в выдыхаемом воздухе может ощущаться запах ацетона.
17.13. Ацетоацетат-основное топливо в некоторых тканях
Основным местом образования ацетоацетата и 3-гидроксибутирата служит печень. Из митохондрий печени эти соединения диффундируют в кровь и переносятся к периферическим тканям. Еще несколько лет назад считалось, что кетоновые тела - это продукты расщепления, не имеющие существенного физиологического значения. Исследования Георга Кэхилла (George Cahill) и других показали, однако, что эти производные ацетил-СоА играют важную роль в энергетическом обмене. Ацетоацетат и 3-гидроксибутират в норме выполняют роль дыхательного топлива и являются количественно важными источниками энергии. Действительно, сердечная мышца и корковый слой почек предпочтительно используют ацетоацетат, а не глюкозу. В противоположность этому глюкоза является главным топливом для мозга у лиц, получающих обильную сбалансированную пищу. Тем не менее при голодании и диабете мозг адаптируется к использованию ацетоацетата. В условиях длительного голодания 75% потребности мозга в топливе удовлетворяется за счет ацетоацетата.
Ацетоацетат может быть активирован путем переноса СоА с сукцинил-СоА в реакции, катализируемой специфической СоА- трансферазой. Ацетоацетил-СоА далее расщепляется тиолазой с образованием двух молекул ацетил-СоА, которые могут затем включиться в цикл трикарбоновых кислот. Печень может снабжать ацетоацетатом другие органы, поскольку в ней отсутствует специфическая СоА-трансфераза.
Ацетоацетат можно рассматривать как водорастворимую транспортируемую форму ацетильных компонентов. Жирные кислоты высвобождаются жировой тканью и превращаются в ацетильные компоненты печенью, которая затем экспортирует их в виде ацетоацетата. Как и можно было предвидеть, ацетоацетат играет и регуляторную роль. Высокое содержание ацетоацетата в крови означает изобилие ацетильных компонентов и приводит к снижению скорости липолиза в жировой ткани.