БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 17. ОБМЕН ЖИРНЫХ КИСЛОТ
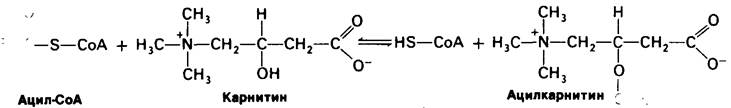
17.7. Карнитин переносит в митохондриальный матрикс активированные жирные кислоты с длинной цепью
Жирные кислоты активируются на наружной митохондриальной мембране, а окисляются в митохондриальном матриксе. Будучи молекулой с длинной цепью, ацил- СоА не может проходить легко через внутреннюю митохондриальную мембрану. Для этого необходим специальный механизм транспорта. Переносчиком активированных жирных кислот с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Ацильная группа переносится с атома серы СоА на гидроксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану. На той стороне этой мембраны, которая обращена к матриксу, ацильная группа переносится обратно на СоА, что термодинамически выгодно, поскольку О-ацильная связь в карнитине обладает высоким потенциалом переноса группы. Эти реакции трансацетилирования катализируются ацил-СоА: карнитин — — ацилтрансферазой.
Можно было думать, что у людей с дефектом данной трансферазы или недостаточностью карнитина должно быть нарушено окисление жирных кислот с длинной цепью. Такое нарушение действительно было обнаружено у идентичных близнецов, у которых с раннего детства наблюдались болезненные мышечные судороги. Боли снимались при голодании, упражнениях и при потреблении богатой жиром пищи; во всех этих трех состояниях основным процессом, обеспечивающим энергию, является окисление жирных кислот. Ферменты гликолиза и гликогенолиза не отличались от нормы. Липолиз триацилглицеролов также был в пределах нормы, о чем свидетельствовало повышение концентрации неэтерифицированных жирных кислот в плазме после голодания. Анализ биопсийного материала мышечной ткани показал, что синтетаза СоА-производных жирных кислот (ацил-СоА) с длинной цепью была полностью активна. Кроме того, нормально протекал метаболизм жирных кислот со средней длиной цепи (С8 и С10). Известно, что для проникновения в митохондриальный матрикс ацил-СоА со средней длиной цепи карнитина не требуется. Описанный случай отчетливо показывает, что нарушение перехода метаболита из одного клеточного компартмента в другой может явиться причиной болезни.
17.8. В каждом цикле окисления жирной кислоты происходит генерирование ацетил-СоА, NАDН и FАDН2
Насыщенный ацил-СоА распадается в результате повторяющейся последовательности из четырех реакций: окисления с участием флавинадениндинуклеотида (FAD) гидратации, окисления с участием NAD+ и тиолиза с участием СоА. Цепь жирной кислоты укорачивается при этом на два атома углерода, и происходит генерирование FADH2, NADH и ацетил-СоА. Дэвид Грин (David Green), Северо Очоа (Severo Ochoa) и Феодор Линен (Feodor Lynen) внесли большой вклад в изучение этой последовательности реакций, назвав его путем fi-окисления.
Первой реакцией в каждом цикле распада ацил-СоА является его окисление ацил-СоА — дегидрогеназой, приводящее к образованию еноил-СоА с транс-двойной связью между С-2 и С-3.
Ацил-СоА + E-FAD → транс-∆2- -Еноил-СоА + E-FADH2.
Интересно отметить, что дегидрирование ацил-СоА очень сходно с дегидрированием сукцината в цикле трикарбоновых кислот. Действительно, первые три реакции в каждом цикле расщепления жирной кислоты близко напоминают последние стадии в цикле трикарбоновых кислот:
Ацил-СоА → Енол-СоА → Гидроксиацил- СоА → Оксоацил-СоА
Сукцинат → Фумарат → Малат → Оксалоацетат.
Следующий этап - гидратация двойной связи между С-2 и С-3 под действием еноил-СоА—гидратазы.
транс-∆2-Еноил-СоА + Н2O → L-3-гидроксиацил-СоА.
Гидратация еноил-СоА стереоспецифична, подобно гидратации фумарата и аконита- та. В результате гидратации транс-∆2-двойной связи образуется только L-изомер 3- гидроксиацил-СоА. Фермент гидратирует также цис-∆2-двойную связь, но в этом случае продуктом реакции является D-изомер. Мы еще вернемся вкратце к этому
моменту при обсуждении окисления ненасыщенных жирных кислот.
Гидратация еноил-СоА предшествует втооой реакции окисления, при которой гидроксильная группа при С-3 превращается в оксогруппу и происходит генерирование NADH. Эта реакция катализируется l-3-ги- дроксиацил-СоА дегидрогеназой, обладающей абсолютной специфичностью в отношении ь-изомера гидроксиацильного субстрата.
L-3-гидроксиацил-СоА + NAD+ ⇄ 3-оксоацил-СоА + NADH + Н+.
В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Последняя стадия представляет собою расщепление 3-оксоацил-СоА тиоловой группой второй молекулы СоА, продуктами которого являются ацетил- СоА и ацил-СоА, укороченные на два атома углерода. Это тиолитическое расщепление катализируется β-кетотиолазой.
3-Оксоацил-СоА + HS—СоА ⇄ Ацетил-СоА + Ацил-СоА.
(n углеродов) (n — 2 углеродов)
Укороченный ацил-СоА подвергается далее следующему циклу окисления, начинающему с реакции, катализируемой ацил-СоА — дегидрогеназой. Бета-кетотиолаза гидроксиацил-дегидрогеназа и еноил-СоА -гидратаза обладают широкой специфичностью в отношении длины ацильной группы.
17.9. При полном окислении пальмитата образуется 129 АТР
Мы можем вычислить, каков выход энергии при окислении жирной кислоты. В каждом цикле реакций ацил-СоА укорачивается на два углерода и образуется по одной молекуле FADH2, NADH и ацетил-СоА.
Сn-ацил-СоА + FAD + NAD+ + Н2O + СоА → Сn-2-ацил-СоА + FADH2 + NADH + Ацетил-СоА + Н+.
Расщепление пальмитоил-СоА (С16-ацил- СоА) требует семи циклов. В седьмом цикле С4-оксоацил-СоА расщепляется путем тиолиза на две молекулы ацетил-СоА. В итоге получаем следующую стехиометрию окисления пальмитоил-СоА:
Пальмитоил-СоА + 7FAD + 7NAD+ + 7СоА + 7Н2O → 8 Ацетил-СоА + 7FADH2 + 7NADH + 7Н+.
При окислении каждого из этих NADH через дыхательную цепь образуются три АТР, тогда как при окислении каждого FADH2 - два АТР, потому что в этом случае электроны поступают в цепь на уровне кофермента Q. Напомним, что окисление ацетил-СоА в цикле трикарбоновых кислот дает 12 АТР. Значит, количество молекул АТР, образованного при окислении пальмитоил-СоА, составит: 14 из 7 FADH2, 21 из 7NADH и 96 из 8 молекул ацетил-СоА, что равно в сумме 131. Две высокоэнергетические фосфатные связи потребляются для активации пальмитата, когда АТР расщепляется на АМР и 2Рi. Таким образом, чистый выход АТР в результате полного окисления пальмитата составляет 129 молекул.
Эффективность запасания энергии при окислении жирных кислот можно измерить, исходя из количества, образованного АТР, и свободной энергии окисления пальмитиновой кислоты до СO2 и Н2O, определяемой калориметрическим путем. Стандартная свободная энергия гидролиза 129 АТР равна — 940 ккал (129 х — 7,3 ккал). Стандартная свободная энергия окисления пальмитата равна — 2340 ккал. Следовательно, эффективность накопления энергии в результате окисления жирных кислот при стандартных условиях составляет около 40%, что близко к этой величине для гликолиза, цикла трикарбоновых кислот и окислительного фосфорилирования.
Таблица 17.2. Основные реакции в окислении жирных кислот
