БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 17. ОБМЕН ЖИРНЫХ КИСЛОТ
17.3. Триацилглицеролы (триглицериды)- высококонцентрироваиные резервы энергии
Триацилглицеролы (триглицериды) в силу своего восстановленного и обезвоженного состояния представляют собою высококонцентрированные резервы метаболической энергии. Выход энергии в результате полного окисления жирных кислот составляет около 9 ккал/г, тогда как для углеводов и белков эта величина равна примерно 4 ккал/г. Такое большое различие в выходе энергии объясняется тем, что жирные кислоты являются значительно более высоковосстановленными соединениями. Кроме того, триацилглицеролы обладают сильно выраженной неполярностью и поэтому резервируются в почти обезвоженной форме, тогда как белки и углеводы значительно более полярны и соответственно более высокогидратированы. Действительно, один грамм сухого гликогена связывает около двух граммов воды. Следовательно, количество энергии, запасенной в одном грамме почти обезвоженного жира, более чем в 6 раз превышает количество энергии, запасенной в одном грамме гидратированного гликогена. Этим и объясняется тот факт, что именно триацилглицеролы, а не гликоген были отобраны в ходе эволюции в качестве основного источника энергии. У человека весом в 70 кг резервы топлива в норме распределяются следующим образом: 100000 ккал в триацилглицеролах, 25 000 ккал в белках (преимущественно в мышечных), 600 ккал в гликогене и 40 ккал в глюкозе. На триацилглицеролы из общего веса тела приходится 11 кг. Если бы это же количество энергии запасалось в виде гликогена, общий вес тела должен был быть на 55 кг больше.
У млекопитающих основным местом накопления триацилглицеролов является цитоплазма жировых клеток. Капли триацилглицерола сливаются, образуя большие глобулы, которые могут занимать большую часть клеточного объема. Жировые клетки специализированы для синтеза и хранения триацилглицеролов, а также для их мобилизации в качестве топливных молекул, способных переноситься кровью к другим тканям.
17.4. Триацилглицеролы гидролизуются липазами, регулируемыми циклическим АМР
Первым этапом в использовании жира как источника энергии является гидролиз триа- цилглицерола под действием липаз. Активность липазы в жировой клетке регулируется гормонами. Адреналин, норадреналин, глюкагон и адренокортикотропный гормон стимулируют аденилатциклазу жировых клеток. Повышенное содержание циклического аденозинмонофосфата (циклического АМР) приводит далее к стимуляции протеинкиназы, которая активирует липазу путем ее фосфорилирования. Таким образом, адреналин, норадреналин, глюкагон и адренокортикотропный гормон вызывают липолиз. Циклический АМР играет роль второго посредника в активации липолиза в жировых клетках, которая аналогична его роли в активации распада гликогена (гл. 16). В отличие от перечисленных гормонов инсулин ингибирует липолиз.
Глицерол, образованный при липолизе, фосфорилируется и восстанавливается до дигидроксиацетонфосфата, который в свою очередь изомеризуется в глицеральдегид-3- фосфат. Последний служит промежуточным продуктом и гликолиза, и глюко- неогенеза. Следовательно, в печени, содержащей соответствующие ферменты, глицерол может превращаться в пируват или глюкозу. Возможен и обратный процесс— восстановление дигидроксиацетонфосфата в глицерол-3-фосфат, который затем гидролизуется фосфатазой с образованием глицерола. Таким образом, глицерол и промежуточные продукты гликолиза легко подвергаются взаимопревращению.
Рис. 17.2. Сканирующая электронная микрофотография жировой клетки
17.5. Жирные кислоты распадаются путем последовательного удаления двухуглеродных фрагментов
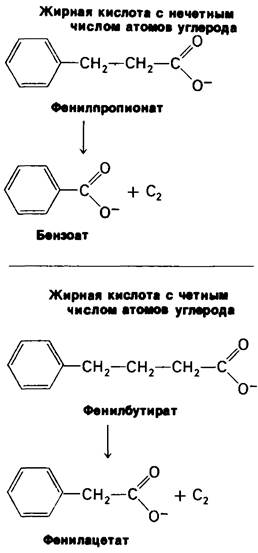
В 1904 г. Франц Кноп (Franz Knoop) сделал решающий вклад в выяснение механизма окисления жирных кислот. Он скармливал собакам жирные кислоты с неразветвленной цепью, в которых ω-углеродный атом был присоединен к фенильной группе. Кноп установил, что при скармливании собакам фенилбутирата в моче обнаруживается производное фенилуксусной кислоты, а при скармливании фенилпропионата - производное бензойной кислоты. Итак, скармливание жирной кислоты с четным числом атомов углерода сопровождалось образованием фенилуксусной кислоты, тогда как скармливание кислоты, содержащей нечетное число атомов углерода, образованием бензойной кислоты (рис. 17.3). На основе этих данных Кноп пришел к выводу, что жирные кислоты расщепляются путем окисления при β-углеродном атоме. Описанные эксперименты представляют собою веху в развитии биохимии, поскольку в них впервые было использовано синтетическое меченое соединение для изучения биохимического механизма. Дейтерий и радиактивные изотопы начали использоваться в биохимии несколькими десятилетиями позднее.
17.6. Окислению жирных кислот предшествует связывание с коферментом А
В 1949 г. Юджин Кеннеди и Альберт Ленинджер (Eugene Kennedy, Albert Lehninger) обнаружили, что окисление жирных кислот происходит в митохондриях. Последующие работы показали, что проникновению жирных кислот в митохондриальный матрикс предшествует их активация. Аденозинтрифосфат (АТР) стимулирует образование тиоэфирной связи между карбоксильной группой жирной кислоты и сульфгидрильной группой СоА. Эта реакция активации происходит в митохондриальной мембране, где она катализируется ацил-СоА — синтетазой (называемой также тиокиназой жирных кислот).
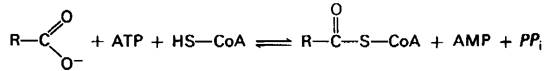
Пауль Берг (Paul Berg) установил, что активация жирной кислоты протекает в два этапа. Сначала жирная кислота реагирует с АТР с образованием ациладенилата. В этом смешанном ангидриде карбоксильная группа жирной кислоты присоединена к фосфорильной группе АМР. Другие две фосфорильные группы ATP-субстрата освобождаются в виде пирофосфата. Далее сульфгидрильная группа СоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-СоА и АМР. Обе эти частичные реакции легкообратимы. Константа равновесия для суммарной реакции близка к 1.
R - СОО-+ СоА + ATR ⇄ Ацил-СоА + АМР + РРi.
Рис. 17.3. Эксперимент Кнопа (Knoop), показывающий, что жирные кислоты распадаются путем отщепления двухуглеродных фрагментов
В ходе описанных превращений одна высокоэнергетическая связь разрушается (связь между PPi и АМР) и одна образуется (тиоэфирная связь в ацил-СоА). Каким образом запускается эта реакция? Ответом на этот вопрос может служить тот факт, что пирофосфат быстро гидролизуется под действием пирофосфатазы.
R - СОО- + СоА + АТР + Н2O → Ацил-СоА + АМР + 2Pi + 2Н+.
В результате вся реакция оказывается необратимой, поскольку потребляются две высокоэнергетические связи, а образуется лишь одна. Мы видим здесь еще один пример повторяющегося в биохимии мотива: многие биосинтетические реакции становятся необратимыми вследствие гидролиза неорганического пирофосфата.
Другой повторяющийся мотив касается реакции активации. Наличие связанного с ферментом ациладенилатного промежуточного продукта присуще не только синтезу ацил-СоА. Aциладенилаты часто образуются при активации карбоксильных групп в биохимических реакциях. Например, именно при помощи такого механизма происходит активация аминокислот для синтеза белка.