БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 17. ОБМЕН ЖИРНЫХ КИСЛОТ
17.20. Жирные кислоты синтезируются у эукариот мультиферментным комплексом
Синтетазы жирных кислот у эукариот в отличие от таковых у бактерий представляют собою совершенно определенный мультиферментный комплекс. Комплекс, выделенный из дрожжей, обладает массой в 2300 кДа и на электронной микрофотографии имеет вид эллипсоида длиной 250 А и поперечным диаметром 210 А (рис. 17.13). Он состоит только из двух видов полипеп- тидпых цепей, каждая из которых кодируется одним геном. Субъединица А (185 кДа) содержит ацилпереносящий белок, конденсирующий фермент и ß-оксоацил — редуктазу, тогда как субъединица В (175 кДа) содержит ацетил-трансацилазу, малонил-трансацилазу, ß-гидроксиацил — дегидратазу и еноилредуктазу. Синтетаза жирных кислот у млекопитающих (400 кДа) также образована двумя видами субъединиц, подобных таковым у дрожжей. На самом деле, многие мультиферментпые комплексы эукариот состоят из полифункциональных белков, в которых различные ферменты ковалентно связаны в единую полипептидную цепь. Преимущество такой организации -возможность координирования синтеза различных ферментов. Кроме того, мульти ферментный комплекс, состоящий из ковалентно соединенных ферментов, является более стабильным, чем комплекс, образованный нековалентными связями.
Рис. 17.13. Электронная микрофотография комплекса синтетазы жирных кислот из дрожжей
Линен (Lynen) предположил, что удлиняющаяся цепь жирной кислоты переносится от АПБ к конденсирующему ферменту и обратно в каждом цикле элонгации. Первая транс локация освобождает место для транспортируемого малой ильного компонента, вторая-происходит на стадии конденсации. Интересно отметить, что аналогичные транслокации имеют место при синтезе белков.
Гибкость и максимальная длина в 20 А фосфопантетеинильного компонента представляются критическими для функции мультиферментного комплекса, поскольку они обеспечивают тесный контакт удлиняющейся цени жирной кислоты с активным центром каждого фермента в комплексе. Для взаимодействия субъединиц фермента с субстратом не требуется их большой структурной перестройки, поскольку сам субстрат на длинном гибком плече может достигнуть каждого активного центра. Напомним, что биотин и липоамид в составе соответствующих мультиферментных комплексов также снабжены длинными гибкими плечами. Организованная структура синтетаз жирных кислот у дрожжей и высших организмов повышает общую эффективность процесса благодаря прямому переносу промежуточных продуктов от одного активного центра к следующему. Реагирующие соединения не разбавляются в цитозоле. Кроме того, им не нужно «находить» друг друга путем случайной диффузии. Еще одним преимуществом такого мультиферментного комплекса является то обстоятельство, что ковалентно связанные промежуточные продукты изолированы и защищены от конкурирующих реакций.
Лиазы-ферменты, катализирующие расщепление связей С—С, С—О или С—N путем элиминирования. В этих реакциях образуется двойная связь.
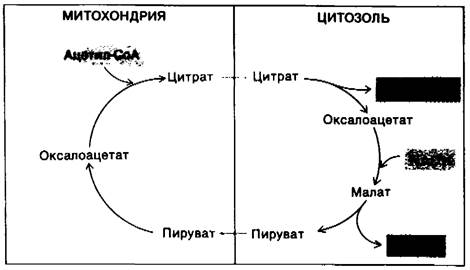
17.21. Цитрат переносит ацетильные группы для синтеза жирных кислот из митохондрий в цитозоль
Синтез пальмитата требует наличия 8 молекул ацетил-СоА, 14 NADPH и 7 АТР. Жирные кислоты синтезируются в цитозоле, тогда как ацетил-СоА образуется из пирувата в митохондриях. Следовательно, для синтеза жирных кислот необходимо, чтобы ацетил-СоА был перенесен из митохондрий в цитозоль. Однако митохондрии непроницаемы для ацетил-СоА. Напомним, что карнитин переносит только длинноцепочечные жирные кислоты. Обход этого барьера для ацетил-СоА осуществляется при помощи цитрата, переносящего ацетильные группы через внутреннюю митохондриальную мембрану. Цитрат образуется в митохондриальном матриксе путем конденсации ацетил-СоА и оксалоацетата. Затем он диффундирует в цитозоль, где расщепляется цитрат-лиазой:
Цитрат + АТР + СоА → Ацетил-СоА + ADP + Рi + Оксалоацетат.
Таким образом, ацетил-СоА и оксалоацетат переносятся из митохондрий в цитозоль с использованием одной молекулы АТР.
17.22. Источники NADPH для синтеза жирных кислот
Оксалоацетат, образованный в результате переноса ацетильной группы в цитозоль, должен быть затем возвращен в митохондрии. Внутренняя митохондриальная мембрана непроницаема для оксалоацетата. Следовательно, необходимы реакции, идущие в обход этого препятствия. Очень важно, что в ходе этих реакций генерируется значительная часть NADPH, необходимого для синтеза жирных кислот. Первая реакция состоит в восстановлении оксалоацетата до малата с участием NADH. Она происходит в цитозоле и катализируется малат-дегидрогеназой.
Оксалоацетат + NADH + Н+⇄ Малат + NAD+.
Вторая реакция - окислительное декарбок- силирование малата NADP+ -зависимой малат-дегидрогеназой (декарбоксилирующей), называемой также «яблочным» ферментом. С этой реакцией мы встречаемся впервые.
Малат + NADP+ → Пируват + СO2 + NADPH.
Образовавшийся пируват легко диффундирует в митохондрии, где он карбоксилируегся в оксалоацетат под действием пируват- карбоксилазы.
Пируват + СO2 + АТР + Н2O → Оксалоацетат + ADP+ + Рi + 2Н+.
Суммируя эти три реакции, получаем
NADP+ + NADH + АТР + Н2O → NADPH + NAD+ + ADP + Pi + H+.
Таким образом, на каждую молекулу ацетил-СоА, которая переходит из митохондрий в цитозоль, образуется одна молекула NADPH. Следовательно, при переходе восьми молекул ацетил-СоА в цитозоль в процессе синтеза пальмитата образуются восемь NADPH. Еще шесть NADPH, требующиеся для этого процесса, генерируются в пентозофосфатном пути.
Рис. 17.14. Ацетил-СоА переносится из митохондрий в цитозоль при сопутствующем превращении NADH в NАDРН в ходе указанной серии реакций