БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 19. ФОТОСИНТЕЗ
19.2. Хлорофиллы представляют собой фоторецепторные молекулы
Майер говорил: «Природа поставила перед собой задачу: как уловить потоки света, льющиеся на землю, и как хранить в твердой форме этот самый неуловимый из всех видов энергии?» Каков механизм улавливания этой самой неуловимой энергии? Первый этап - поглощение света фоторецепторной молекулой. Основным фоторецептором в хлоропластах зеленых растений служит хлорофилл а, замещенный тетрапиррол. Четыре атома азота пиррольных колец координированы с атомом магния. Таким образом, хлорофилл представляет собою магнийпорфирин, тогда как гем является железопорфирином. Порфи- риновая кольцевая структура хлорофилла отличается от структуры гема по нескольким параметрам: 1) одно из пиррольных колец частично восстановлено; 2) с одним из пиррольных колец слито циклопентаноновое кольцо; 3) в хлорофилле обе кислотные боковые цепи этерифицированы, тогда как в геме они свободны. Одна из кислотных цепей в хлорофилле представляет собою метиловый эфир, вторая- эфир фитола (С20С39ОН). Этот многоатомный спирт состоит из четырех изопреновых единиц, которые сообщают ему высокую гидрофобность. Хлорофилл b отличается от хлорофилла а природой заместителя при одном из пиррольных колец: в хлорофилле b это-формильная группа вместо метильной группы в хлорофилле а.
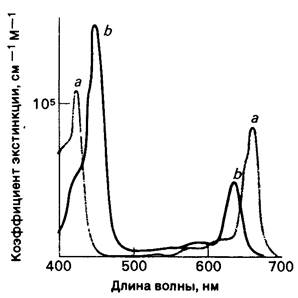
Указанные хлорофиллы - очень эффективные фоторецепторы, поскольку они содержат сеть чередующихся одинарных и двойных связей. Другими словами, они представляют собой полиены. Они сильно поглощают в видимой части спектра, соответствующей максимуму достигающего Земли солнечного излучения. Пиковые величины коэффициентов экстинкции хлорофиллов а и b превышают 1015см-1м-1 и находятся в ряду самых высоких значений этих коэффициентов, известных для органических соединений.
Спектры поглощения хлорофиллов а и b различны (рис. 19.4). Свет, который не поглощается в заметной степени хлорофиллом а, например, при 460 нм, улавливается хлорофиллом b, который обладает интенсивным поглощением именно при этой длине волны. Таким образом, два вида хлорофилла дополняют друг друга в поглощении солнечного света. В то же время имеется большая область спектра, от 500 до 600 нм, в которой поглощение света происходит относительно слабо. Большинство растений, однако, не нуждается в улавливании света в этой области спектра, так как для них достаточно света, поглощаемого в синей и красной частях спектра.
Рис. 19.4. Спектры поглощения хлорофиллов а и b
19.3. Первичные события фотосинтеза происходят в высокоорганизованной системе мембран
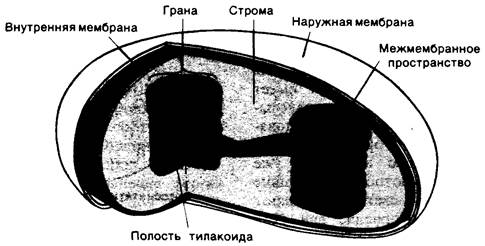
Хлoponласты, органеллы фотосинтеза, обычно достигают в длину 5 мкм. Подобно митохондрии, хлоропласт имеет наружную и внутреннюю мембраны с межмембранным пространством между ними (рис. 19.5). Внутренняя мембрана окружает строму, содержащую растворимые ферменты и мембранные структуры, называемые тилакоидами, которые представляют собою уплощенные мешочки. Стопки таких мешочков образуют граны. Различные граны соединяются между собою участками мембраны, называемыми stromalamellae. Мембраны тилакоидов отделяют полость тилакоидов от полости стромы. Таким образом, в хлоропластах имеются три вида мембран (наружная, внутренняя и тилакоидная) и три отдельные полости (межмембранное пространство, полость стромы и полость тилакоидов). В развивающихся хлоропластах тилакоиды возникают из впячиваний во внутренней мембране, и, следовательно, они аналогичны митохондриальным кристам.
Рис. 19.5. Схематическое изображение хлоропласта
Мембраны тилакоидов содержат молекулы хлорофилла и другие компоненты преобразующего энергию механизма. Количество липидов и белков примерно одинаково. Липидный состав совершенно определенный: около 40% общего количества липидов составляют галактолипиды, 4%-сулъфолипиды, тогда как на фосфолипиды приходится всего 10%. Мембрана тилакоидов, подобно внутренней митохондриальной мембране, непроницаема для большинства молекул и ионов. В строме присутствуют растворимые ферменты, которые используют синтезируемые тилакоидами NADPH и АТР для превращения СО2 в сахар. В хлоропластах имеются собственная ДНК (разд. 29.9) и механизм для синтеза РНК и белка. Таким образом, хлоропласт представляет собою органеллу, обладающую значительной автономией. Внутренняя мембрана хлоропласта, содержащая транслокаторы для ряда соединений, таких, как АТР и дикарбоновые кислоты, служит местом взаимодействия между хлоропластом и остальной частью клетки. Наружная мембрана хлоропласта, как и таковая митохондрии, обладает высокой проницаемостью для небольших молекул и ионов.
19.4. Фотосинтетическая единица: фотоны стекаются в реакционный центр
При измерении скорости фотосинтеза как функции интенсивности освещения оказывается, что она линейно возрастает при низких значениях интенсивности и достигает насыщающей величины при ее высоких значениях (рис. 19.6). Насыщающая величина отмечается в условиях сильного освещения, поскольку химические реакции использования поглощенных фотонов лимитируют скорость процесса. Таким образом, фотосинтез можно подразделить на световые и темновые реакции. Как будет вскоре указано, световые реакции генерируют NADPH и АТР, а в темновых реакциях эти богатые энергией молекулы используются для восстановления CO2.
Рис. 19.6. Скорость фотосинтеза достигает лимитирующей величины, когда интенсивность освещения достаточна для возбуждения только небольшой доли молекул хлорофилла

В 1932 г. Роберт Эмерсон и Уильям Арнольд (Robert Emerson, William Arnold) измерили выход кислорода при фотосинтезе в условиях, когда на клетки Chlorella воздействовали световыми вспышками длительностью в несколько микросекунд. Они предполагали, что выход фотосинтеза на вспышку будет увеличиваться с усилением интенсивности вспышки до тех пор, пока каждая молекула хлорофилла не поглотит фотон, который будет затем использован в темновых реакциях. Результат эксперимента был совершенно неожиданным: насыщающая световая вспышка приводила к образованию только одной молекулы O2 на каждые 2500 молекул хлорофилла.
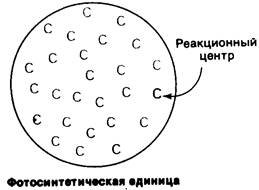
В результате этих исследований была постулирована концепция фотосинтетической единицы. Ганс Гаффрон (Hans Gafïron) предположил, что свет поглощается сотнями молекул хлорофилла, которые затем переносят свою энергию возбуждения к тому месту, где протекают химические реакции (рис. 19.7). Это место называется реакционным центром. Таким образом, функция большинства молекул хлорофилла в фотосинтетической единице состоит в поглощении света. Только малая доля хлорофиллов, те, которые локализованы в реакционных центрах, участвует в преобразовании света в химическую энергию. Хлорофиллы в реакционном центре химически идентичны другим хлорофиллам фотосинтетической единицы, но обладают особыми свойствами, обусловленными их особым окружением. Одно из различий состоит в том, что энергетический уровень возбужденного состояния хлорофиллов реакционного центра ниже, чем у других хлорофиллов, и они поэтому способны улавливать энергию. Энергия, поглощенная молекулами хлорофилла, перемещается по фотосинтетической единице, пока не достигнет хлорофилла реакционного центра. Перенос энергии к реакционному центру идет очень быстро, занимая менее 10-10 с.
Рис. 19.7. Схема фотосинтетической единицы. Молекулы хлорофилла антенн (обозначенные синими буквами С) переносят свою энергию возбуждения на специализированный хлорофилл в реакционном центре (обозначенный красной буквой С)