БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 19. ФОТОСИНТЕЗ
19.15. СO2 реагирует с рибулозобисфосфатом с образованием двух фосфоглицератов
Радиохроматограмма, получаемая после 60 с освещения, оказалась настолько сложной (рис, 19.19), что на ней невозможно было отличить самый ранний промежуточный продукт в процессе фиксации СО2. Однако после освещения в течение всего лишь 5 с картина была значительно проще: на хроматограмме присутствовало только одно явное радиоактивное пятно, которое оказалось 3-фосфоглицератом.
Рис. 19.18. Электронная микрофотография мембран тилакоидов шпината, полученная методом замораживания - скалывания. Видна правильная решетка трансмембранных частиц
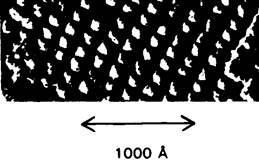
Рис. 19.19. Радиохроматограммы освещенных суспензий водоросли через 5 и 60 с после введения в систему СO2

Образование 3-фосфоглицерата в качестве первого обнаруживаемого радиоактивного промежуточного продукта наводило на мысль, что акцептором СО2 служит двухуглеродное соединение. Оказалось, что это не так. Действительная последовательность реакций гораздо сложнее:
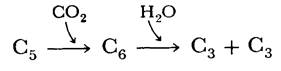
Молекула СО2 конденсируется с рибулозо-1,5-бисдифосфатом с образованием короткоживущего шестиуглеродного соединения, которое быстро гидролизуется до двух молекул 3-фосфоглицерата. Суммарная реакция носит высокоэкзергонический характер (∆G0 = 12,4 ккал/моль).
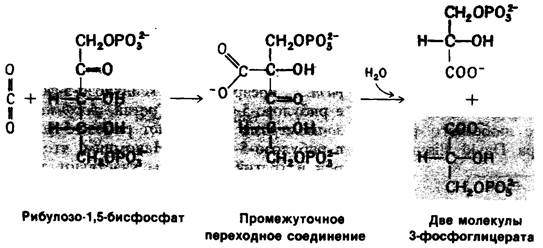
Реакция катализируется рибулозо-1,5-бисфосфат—карбоксилазой, которая локализована на обращенной к строме поверхности мембран тилакоидов. Этим ферментом очень богаты хлоропласты; на его долю приходится более 16% общего содержания в них белка. Рибулозо-1,5-бисфосфат—карбоксилаза, вероятно, является белком, содержащимся в биосфере в наибольшем количестве. Он состоит из восьми больших (55 кДа) и восьми малых (15 кДа) субъединиц, расположенных в виде двух слоев (рис. 19.20). Большие субъединицы каталитически активны и в отсутствие малых субъединиц, которые выполняют регуляторную роль. Фермент является в одно и то же время и оксигеназой (разд. 19.20), и карбоксилазой.
Рис. 19.20. Схематическое изображение структуры рибулозо-1,5-бис- фосфат—карбоксилазы

19.16. Образование фруктозо-6-фосфата и регенерирование рибулозо-1,5-бисфосфата
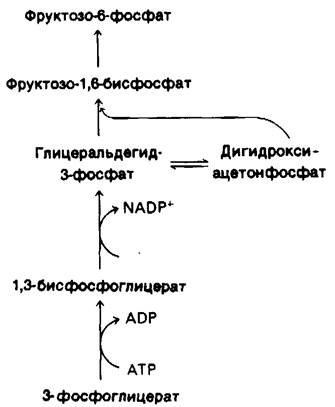
Этапы превращения 3-фосфоглицерата во фруктозо-6-фосфат (рис. 19.21) подобны таковым в пути глюконеогенеза (разд. 15.13) с той лишь разницей, что глицеральдегид-3- фосфат—дегидрогеназа хлоропластов специфична в отношении NADPH, а не NАDН. В результате этих реакций СO2 переходит на уровень гексозы. Остается лишь регенерировать рибулозобисфосфат, акцептор СO2 на первой темновой стадии. Проблема сводится к образованию пятиуглеродного сахара из шестиуглеродного и трехуглеродного сахаров. Оно осуществляется в ходе реакций, катализируемых транскетолазой и альдолазой. Транскетолаза участвует также в превращениях по пентозофосфатному пути (разд. 15.4).
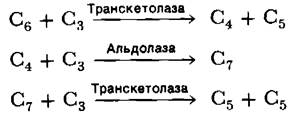
Напомним, что транскетолаза, фермент, использующий в качестве простетической группы тиаминпирофосфат, переносит двухуглеродный фрагмент (СН2ОН—СО—) с кетозы на альдозу. Аль- долаза осуществляет альдольную конденсацию между дигидроксиацетонфосфатом и альдегидом. Фермент этот высокоспецифичен для дигидроксиацетонфосфата, но реагирует с множеством альдегидов. К специфическим реакциям, катализируемым транскетолазой и альдолазой в цикле Кальвина, относятся следующие:

Рис. 19.21. Путь превращения 3-фосфо- глицерата во фруктозо-6-фосфат в хлоропластах
Для темновых реакций фотосинтеза требуются четыре дополнительных фермента. Один из них, фосфатаза, гидролизует седо- гептулозо-1,7-бисфосфат до седогептулозо-7-фосфата. Второй, фосфопентозо-эпиме- раза, превращает ксилулозо-5-фосфат в рибулозо-5-фосфат. Третий, фосфопентозо-изомераза, превращает рибозо-5-фосфат в рибулозо-5-фосфат. Напомним, что второй и третий ферменты участвуют также в превращениях по пентозофосфатному пути (разд. 15.4). Суммируя вышеуказанные реакции, получаем
Фруктозо-6-фосфат + 2-глицеральдегид-З- фосфат + Дигидроксиацетонфосфат → 3 Рибулозо-5-фосфат.
Наконец, четвертый фермент, фосфорибулозокиназа, катализирует фосфорилирование рибулозо-5-фосфата с регенерированием ри- булозо-1,5-бисфосфата, акцептора СO2.
Рибулозо-5-фосфат + АТР → Рибулозо-1,5-бисфосфат + ADP + Н+.
Эта группа реакций носит название цикла Кальвина (рис. 19.22).
Рис. 19.22. Цикл Кальвина. Детальный путь образования рибулозо-5- фосфата из трехуглеродных и шестиуглеродных сахаров на этой схеме не показан
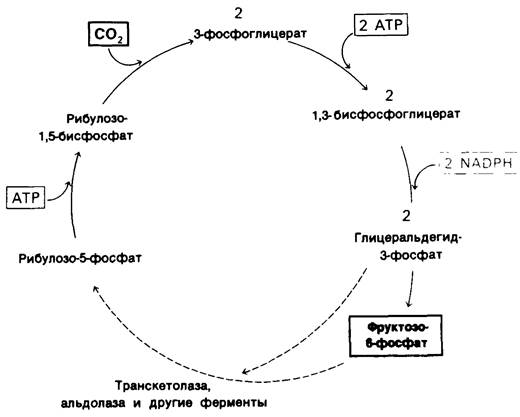
19.17. Три АТР и два NADPH доводят СO2 до уровня гексозы
Каков расход энергии для синтеза гексозы? Требуется шесть оборотов цикла Кальвина, поскольку в каждом из них восстанавливается один атом углерода (рис. 19.22). 12 АТР расходуются на фосфорилирование 12 молекул 3-фосфоглицерата в 1,3-бисфосфо- глицерат и 12 NADPH потребляются при восстановлении 12 молекул 1,3-бисфосфо- глицерата в глицеральдегид-3-фосфат. Еще шесть АТР расходуется на регенерирование рибулозо-1,5-бисфосфата.
Теперь напишем уравнение равновесия для суммарной реакции цикла Кальвина:
6СO2 + 18АТР + 12NADPH + 12Н2O → С6Н12O6 + 18ADP + 18Pi + 12NADP+ + 6Н+.
Таким образом, при превращении СO2 в гексозу (глюкозу или фруктозу) потребляются три молекулы АТР и две молекулы NADPH. Эффективность фотосинтеза может быть измерена следующим образом:
1. ∆G0 для восстановления СO2 до уровня гексозы составляет 114 ккал/моль.
2. При восстановлении NADP+ происходит перенос двух электронов. Следовательно, образование двух NADPH требует засасывания фотосистемой 1 четырех фотонов. Электроны, отдаваемые фотосистемой I, восполняются фотосистемой II, которая должна поглотить эквивалентное количество протонов. Значит, для генерирования, требуемого NADPH необходимы восемь фотонов. Одновременно образуются три молекулы АТР, которые нужны для превращения СO2 в гексозу.
3. Содержание энергии в моле фотонов с длиной волны 600 нм равно 47,6 ккал, и, следовательно, приток энергии от восьми молей фотонов составит 381 ккал. Таким образом, общая эффективность фотосинтеза при стандартных условиях составляет минимум 114/381, или 30%.