БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 19. ФОТОСИНТЕЗ
19.18. Регуляция цикла Кальвина
Существует ряд регуляторных механизмов, обеспечивающих функционирование цикла Кальвина лишь при условии образования АТР и NADPH в ходе световых реакций фотосинтеза. Стадией, лимитирующей скорость, в цикле Кальвина является карбоксилирование рибулозо-1,5-бисфосфата с образованием двух молекул 3-фосфоглицерата. Активность рибулозо-1,5-бисфосфат—карбоксилазы значительно увеличивается при освещении; это увеличение осуществляется тремя путями:
1. NADPH, образуемый фотосистемой I, служит аллостерическим активатором карбоксил азы.
2. Скорость ферментативной реакции значительно возрастает при повышении pH с 7 до 9. Индуцируемое светом закисление полости тилакоида приводит к защелачиванию стромы, которое активирует карбоксилазу.
3. Карбоксилаза активируется Mg2 +. Напомним, что Mg2 + высвобождается в строму при засасывании протонов в полость ти- лакоидов под действием освещения (разд. 19.13).
19.19. У тропических растений функционирует С4-путь, ускоряющий фотосинтез путем концентрирования СО2
У тропических растений, таких, как сахарный тростник, имеется дополнительный путь транспорта СO2 к месту локализации цикла Кальвина в фотосинтезирующих клетках. Источником первых сведений о существовании этого пути явились исследования, показавшие, что радиоактивность импульсной метки 14СO2 первоначально появляется в четырехуглеродных соединениях малате и аспартате, а не в 3-фосфоглицера- те. Смысл данного пути, раскрытого Хэчем и Слэком, состоит в том, что С4-соединения переносят СO2от мезофилъных клеток, соприкасающихся с воздухом, к клеткам обкладки сосудистого пучка, которые служат основным местом локализации фотосинтеза (рис. 19.23). Декарбоксилирование С4-соединения в клетках обкладки сосудистого пучка поддерживает высокую концентрацию СО2 в месте локализации цикла Кальвина. Образовавшееся трехуглероднос соединение возвращается в мезофильную клетку для следующего цикла карбоксилирования.
Рис. 19.23. Схематическое изображение основных этапов С4-пути
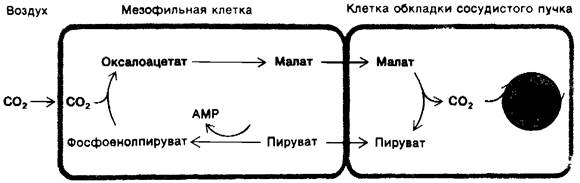
Этот С4-путь (рис. 19.23) для транспорта СO2 начинается в мезофильной клетке с конденсации СO2 и фосфоенолпирувата, приводящей к образованию оксалоацетита, реакции, катализируемой фосфоенолпиру- ват-карбоксилазой. У некоторых видов оксалоацетат превращается в малат под действием NADP+-зависимой малатдегидрогеназы. Малат поступает в клетки обкладки сосудистого пучка и декарбоксилируется в хлоропластах NADP+-зависимой малат- дегидрогеназой. Высвободившийся СO2 включается в цикл Кальвина обычным путем, конденсируясь с рибулозо-1,5-бисфосфатом. Образовавшийся при декарбоксили- ровании пируват возвращается в мезофильную клетку. Наконец, происходит регенерирование фосфоенолпирувата, в ходе реакции пируватас АТР и Рi. Эта единственная реакция, катализируемая пируват-Pi-дикиназой, запускается последующим гидролизом РРi. Суммарная реакция описанного С4-пути выглядит следующим образом:
СO2 (в мезофильной клетке) + АТР + Н2O → СO2 (в клетке обкладки сосудистого пучка) + АМР + 2Рi.
Таким образом, при транспорте СO2 в хлоропласты клеток обкладки сосудистого пучка потребляются две высокоэнергетические фосфатные связи.
В случае одновременного функционирования С4-пути и цикла Кальвина суммарная реакция приобретает следующий вид:
6СO2 + ЗОАТР + 12NADPH + 12Н2O - С6Н12O6 + 30ADP + З0Рi + 12NADP+ + 18Н+.
Отметим, что при поступлении СO2 в цикл Кальвина по С4-пути на одну молекулу образующейся гексозы потребляется 30 АТР, а при отсутствии С4-пути-18 АТР. Высокая концентрация СО2 в клетках обкладки сосудистого пучка растений, у которых функционирует С4-путь, обусловленная расходом дополнительных 12 АТР, является критической для высокой скорости фотосинтеза в этих растениях, поскольку в условиях избытка света СO2 служит лимитирующим фактором. Высокая концентрация СO2 также сводит к минимуму потерю энергии, вызываемую фотодыханием, процессом, который описывается в следующей главе.
19.20. Гликолат - основной субстрат при фотодыханни
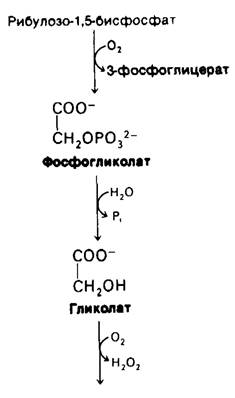
Освещаемые растения поглощают O2 и выделяют СO2 в ходе процесса, называемого фотодыханием, которое отличается от митохондриального дыхания. Биологическая роль этого процесса представляет собою загадку. Гликолат, основной субстрат фотодыхания, происходит из фосфогликолата, образующегося при оксигенировании рибу- лозо-1,5-бисфосфата (рис. 19.24). Реакция катализируется рибулозо-1,5-бисфосфат-карбоксилазой, которая является одновременно и оксигеназой, и карбоксилазой. На самом деле, оксигенирование и карбоксили- рование представляют собою конкурирующие реакции, использующие один и тот же активный центр. Образующийся при этом фосфогликолат гидролизуется под действием специфической фосфатазы до гликолата. Последующий обмен гликолата протекает в пероксисомах (называемых также микротельцами) (рис. 19.25). Гликолат окисляется гликолатоксидазой в глиоксилат. Н2O2, образующийся при этой реакции, расщепляется каталазой на Н2O и O2. Далее глиоксилат подвергается трансаминированию, давая глицин. В митохондриях может происходить образование серина из двух молекул глицина.
Рис. 19.24. Образование и распад гликолата
Фотодыхание представляется непроизводительным процессом, при котором органический углерод превращается в СO2 без образования АТР и NADH или другой видимой «прибыли». Растения, у которых отсутствует С4-путь, теряют при фотодыхании от 25 до 50% фиксированного ими углерода. В противоположность им у тропических растении, обладающих С4-путем, уровень фотодыхания низок, потому что оксигенирование рибулозо-1,5-бисфосфата конкурентно подавляется высокой концентрацией СО2 в клетках обкладки сосудистого пучка. Оксигеназная активность рибулозо-1,5-бисфосфат—карбоксилазы при повышении температуры возрастает быстрее, чем ее карбоксилазная активность. Следовательно, С4-путь играет особенно важную роль в снижении активности фотодыхания до минимума при высоких температурах, Географическое распределение С4-растений (обладающих С4-путем) и С3-растений (у которых этот путь отсутствует) можно теперь понять на молекулярном уровне. С4-растения имеют преимущества в условиях высокой температуры окружающей среды и интенсивного освещения, и поэтому они преобладают в тропиках. С3-растения, у которых на молекулу образующейся гексозы потребляется в отсутствие фотодыхания 18 АТР (по сравнению с 30 АТР у С4-растений), более эффективно функционируют при температурах ниже 28°С, и они преобладают в условиях умеренного климата.
Рис. 19.25. Электронная микрофотография пероксисомы растительной клетки