БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ III. БИОСИНТЕЗ ПРЕДШЕСТВЕННИКОВ МАКРОМОЛЕКУЛ
ГЛАВА 20. БИОСИНТЕЗ МЕМБРАННЫХ ЛИПИДОВ И СТЕРОИДНЫХ ГОРМОНОВ
20.15. Синтез холестерола в печени подавляется холестеролом, поступающим с пищей
Холестерол может поступать с пищей или синтезироваться de novo. Основное место синтеза холестерола у млекопитающих — печень. Кроме того, довольно значительное количество холестерола синтезируется в кишечнике. В норме в организме взрослого человека, получающего обедненный холестеролом рацион, синтезируется около 80 мг холестерола в сутки. Скорость образования холестерола в этих органах в большой степени зависит от количества холестерола, потребляемого с пищей. Этот регуляторный механизм обратной связи действует путем изменения активности З-гидрокси-З-метилглутарил-СоА—редуктазы. Как говорилось выше, данный фермент катализирует образование мевалоната-реакцию, с которой начинается путь биосинтеза холестерола. Поступающий с пищей холестерол подавляет синтез редуктазы в печени и приводит к инактивации уже существующих молекул фермента.
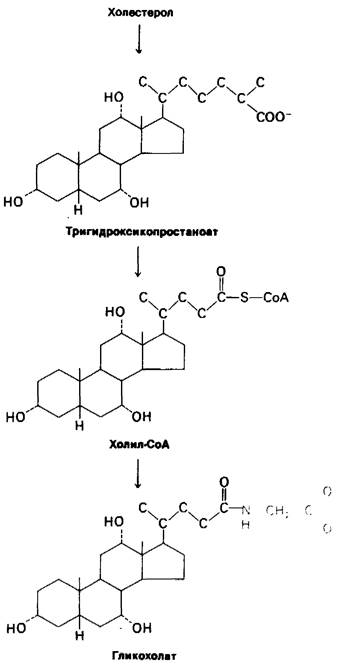
Рис. 20.15. Синтез гликохолата -основной желчной кислоты
20.16. Холестерол и другие липиды переносятся к органам-мишеням с помощью различных липопротеинов
Холестерол, триацилглицеролы и другие липиды транспортируются жидкостями организма с помощью ряда липопротеинов, которые классифицируют по плотности (в порядке возрастания) (табл. 20.1): хиломикроны, липопротеины очень низкой плотности /ЛОНП), липопротеины низкой плотности (ЛНП) и липопротеины высокой плотности (ЛВП). Эти липопротеины состоят из гидрофобного липидного ядра, окруженного полярными липидами и оболочкой из апобелков. К настоящему времени выделено и охарактеризовано восемь типов апобелков: апо-А-I, апо-А-II, апо-В, апо-С-I, апо-С-II, апо-С-III, апо-D и апо-Е. Эти комплексы солюбилизируют сильногидрофобные липиды. Кроме того, белки, входящие в состав этих комплексов, несут сигналы, регулирующие поступление определенных липидов в специфические ткани- мишени и выход липидов из этих тканей.
Таблица 20.1. Липопротеины плазмы

Липопротеины плазмы синтезируются и секретируются печенью и кишечником. Хиломикроны, самые крупные липопротеины, переносят триацилглицеролы, холестерол и другие липиды пищи из кишечника в жировую ткань и печень. Они имеют очень низкую плотность (<0, 94 г/см3), так как богаты триацилглицеролами, и содержат менее 2% белка. Триацилглицеролы в составе хиломикронов за несколько минут гидролизуются липазами в капиллярах жировой ткани и других периферических тканей. Обогащенные холестеролом остатки хиломикронов, так называемые остаточные частицы, поглощаются печенью. Липопротеины очень низкой плотности синтезируются главным образом в печени. ЛОНП доставляют новосинтезированные в организме триацилглицеролы к жировой ткани. Tе ЛОНП, которые не попадают в жировую ткань, превращаются в липопротеины низкой плотности, богатые эфирами холестерола. Большая часть холестерола в ЛНП находится в виде эфира линолевой кислоты- полиеновой жирной кислоты. Роль ЛНП заключается в переносе холестерола к периферическим тканям и регуляции синтеза холестерола de novo в этих тканях; этот аспект мы рассмотрим ниже. Липопротеины высокой плотности синтезируются в печени; они богаты фосфолипидами и холестеролом. Одна из функций ЛВП состоит в переносе холестерола от периферических тканей к печени.
20.17. Рецепторы липопротеинов низкой плотности играют ключевую роль в регуляции метаболизма холестерола
Холестерол - компонент всех клеточных мембран эукариот. Он необходим для роста и существования клеток высших организмов. Однако избыток холестерола может оказаться губительным для организма из-за атеросклероза-отложения бляшек эфиров холестерола. Очевидно, что метаболизм холестерола должен тонко регулироваться. Механизмы регуляции в печени-основном месте синтеза холестерола-уже обсуждались: холестерол, поступающий с пищей, снижает активность и количество З-гидрокси-З-метилглутарил- СоА—редуктазы - фермента, катализирующего решающий этап биосинтеза. Исследования человеческих фибробластов в культуре, проведенные Майклом Брауном и Джозефом Голдстайном (Michael Brown, Joseph Goldstein), пролили свет на регуляцию метаболизма холестерола в других клетках. Как правило, клетки, находящиеся вне печени и кишечника, получают холестерол из сыворотки крови, а не синтезируют его de novo. Точнее, основной источник холестерола-липопротеины низкой плотности. Поглощение клеткой холестерола в составе ЛНП включает следующие этапы:
1. ЛНП связывается со специфическим рецептором на плазматической мембране клеток вне печени. Рецепторы для ЛНП располагаются в особых участках, которые называются «окаймленные рецепторные ямки» (coated pits) и содержат клатрин (разд. 29.32).
2. Комплекс рецептор - ЛНП проникает в клетку путем эндоцитоза, т. е. клеточная мембрана вблизи комплекса втягивается и затем ее края сливаются, образуя эндоцитозный пузырек (рис. 20.16).
Рис. 20.16. Эндоцитоз ЛНП, связанных с соответствующим рецептором на поверхности фибробластов человека в культуре (ЛНП сделаны видимыми путем присоединения ферритина). А - электронная микрофотография; виден ЛНП-ферритин (темные точки), связанный с окаймленной рецепторной ямкой на клеточной поверхности; Б-этот участок в ходе эндоцитоза инвагинируется и образует эндоцитозный пузырек

3. Пузырьки, содержащие ЛНП, сливаются с лизосомами, в которых имеется множество самых разнообразных гидролитических ферментов. Белковый компонент ЛНП гидролизуется до свободных аминокислот. Эфиры холестерола в составе ЛНП гидролизуются кислой липазой лизосом.
4. Высвободившийся неэтерифицированньй холестерол может быть использован в биосинтезе мембран. Кроме того, он может реэтерифицироваться для запасания в клетке. Холестерол активирует ацил-СоА: холестерол—ацилтрансферазу, фермент, катализирующий эту реакцию. Реэтерифицированный холестерол связан преимущественно с олеатом или пальмитолеатом - моноеновымикислотами (т. е. жирными кислотами с одной ненасыщенной связью) в отличие от эфиров холестерола в составе ЛНП, которые обогащены полиеновой линолевой кислотой.
Содержание холестерола в клетках, в которых активно идет процесс усвоения ЛНП, регулируется двумя путями. Во- первых, высвобождающийся холестерол подавляет образование З-гидрокси-З-метил- глутарил-СоА—редуктазы, ингибируя таким образом синтез холестерола de novo. Во-вторых, сам рецептор ЛНП регулируется по механизму обратной связи. В фибробластах время полужизни рецептора ЛНП-около суток. Если в клетках имеется избыток холестерола, новые рецепторы ЛНП не синтезируются, и тем самым блокируется поступление новых порций холестерола из ЛНП плазмы крови.