БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 25. ИНФОРМАЦИЯ РНК И ТРАНСКРИПЦИЯ
25.5. Опыты по гибридизации показали, что информационная РНК комплементарна кодирующей ее ДНК-матрице
В 1961 г. Сол Спигелман (Sol Spigelman) разработал новый метод, названный гибридизацией. Он должен был дать ответ на следующий вопрос: комплементарна ли последовательность оснований РНК, синтезированной после заражения фагом Т2, последовательности оснований ДНК фага Т2? Из работы Джулиуса Мармура и Пола Доти (Julius Marmur, Paul Doty) было известно, что при нагревании двухспиральной ДНК выше температуры плавления она переходит в одноцепочечную форму. При медленном охлаждении раствора эти цепи реассоциируют и образуют двухспиральную структуру, обладающую биологической активностью. Мармур и Доти обнаружили также, что двухспиральные молекулы образуются только в том случае, если цепи ДНК происходят из организмов одного вида или близкородственных видов. Это наблюдение подсказало Спигелману, что в смеси одноцепочечной ДНК и РНК должны образовываться гибриды ДНК-РНК, если их последовательности оснований комплементарны (рис. 25.5). Схема эксперимента была следующей:
1. РНК, синтезированную после заражения Е. coli фагом Т2 (Т2-мРНК), метили изотопом 32Р. В другом эксперименте ДНК фага Т2 (Т2-ДНК) пометили изотопом 3Н.
2. Смесь Т2-мРНК и Т2-ДНК нагревали до 100°С. В результате двухспиральная ДНК расплавлялась и переходила в одноцепочечную форму. Этот раствор, содержащий одноцепочечные РНК и ДНК, медленно охлаждали до комнатной температуры.
3. Охлажденную смесь анализировали методом центрифугирования в градиенте плотности. Образцы центрифугировали несколько дней в бакет-роторе. Затем пластиковые пробирки с образцами протыкали снизу и собирали фракции по каплям для дальнейшего анализа.
Рис. 25.5. Если РНК и ДНК имеют комплементарные последовательности, может образоваться гибрид РНК—ДНК

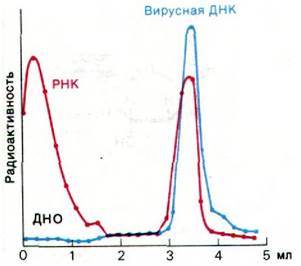
В результате были обнаружены три полосы (рис. 25.6). Полоса с наибольшей плотностью соответствовала одноцепочечной РНК. Вторая полоса соответствовала двухспиральной ДНК. Третья располагалась вблизи от полосы ДНК и состояла из двухцепочечных гибридных молекул ДНК—РНК. Итак, Т2-мРНК образовывала гибрид с Т2-ДНК. В отличие от этого Т2-РНК не гибридизовалась с ДНК многих бактерий и неродственных вирусов, даже если их нуклеотидный состав был подобен Т2-ДНК. Последующие эксперименты показали, что фракция мРНК из незараженных клеток гибридизуется с ДНК именно того организма, из которого она была выделена, но не с ДНК неродственных организмов. Эти убедительные эксперименты продемонстрировали, что последовательность оснований мРНК комплементарна последовательности ДНК-матрицы. К тому же был разработан мощный метод, с помощью которого можно было исследовать поток генетической информации в клетках и выяснять, сходны ли две молекулы нуклеиновой кислоты.
Рис. 25.6. РНК, образовавшаяся после заражения E. coli фагом Т2, комплементарна вирусной ДНК. В этих экспериментах по гибридизации РНК метили 32Р, а ДНК фага Т2 - 3Н. Распределение радиоактивности в градиенте плотности хлористого цезия показывает, что большая часть РНК, синтезированной после заражения, попадает в одну полосу с ДНК фага Т2
25.6. Рибосомные РНК и транспортные РНК также синтезируются на ДНК-матрице
Метод гибридизации был использован затем, чтобы выяснить, синтезируются ли рРНК и тРНК также на ДНК-матрицах. Образование гибридов РНК—ДНК выявляли с помощью фильтров, а не центрифугированием в градиенте плотности, так как этот метод проще, чувствительнее и требует меньше времени. Одноцепочечные РНК проходят через нитроцеллюлозный фильтр, а двухспиральные ДНК и гибриды РНК—ДНК задерживаются на фильтре. РНК Е. coli пометили изотопом 32Р и смешали с немеченой ДНК Е. coli. Эту смесь нагревали, медленно охлаждали и затем отфильтровывали через нитроцеллюлозу. Радиоактивность, задержанную на фильтре, просчитывали. Результаты экспериментов не вызывали сомнений: гибриды РНК—ДНК образовывались со всеми рРНК (5S, 16S и 23S) и тРНК. Отсюда следовало, что в геноме Е. coli имеются последовательности, комплементарные этим молекулам РНК.
25.7. Все клеточные РНК синтезирует РНК-полимеразa
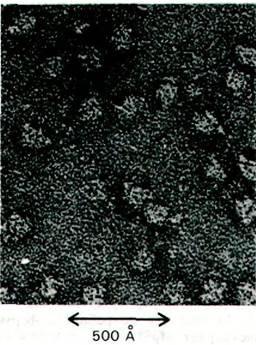
Концепция мРНК стимулировала поиски фермента, который синтезирует РНК в соответствии с последовательностью ДНК- матрицы. Стратегия эксперимента была такой же, как и при поиске ДНК-полимеразы I. В 1960г. Джерард Хёрвиц и Сэмюэл Вейсс (Jerard Hurwitz, Samuel Weiss) независимо открыли такой фермент. Они назвали его РНК-полимеразой. Ферменту из клеток Е. соli (рис. 25.7) нужны были для синтеза РНК следующие компоненты.
Рис. 25.7. Электронная микрофотография РНК-полимеразы E. coli
1. Матрица. Предпочтительная матрица - двухцепочечная ДНК. Одноцепочечная ДНК также может служить матрицей. Ни РНК (как одноцепочечная, так и двухцепочечная), ни гибриды РНК—ДНК не могут служить эффективными матрицами.
2. Активированные предшественники. Необходимы все четыре рибонуклеозидтри- фосфата - АТР, GTP, UTP и СТР.
3. Двухвалентные ионы металлов. Фермент активен в присутствии Mg2+или Mn2+. In vivo эту потребность фермента удовлетворяет Mg2+.
РНК-полимераза катализирует инициацию и элонгацию цепей РНК. Фермент катализирует следующую реакцию:
![]()
Синтез РНК во многих отношениях подобен синтезу ДНК (рис. 25.8). Во- первых, как будет вскоре показано более подробно, синтез идет в направлении 5' → 3'. Во-вторых, по-видимому, механизм элонгации сходен. Происходит нуклеофильная атака внутреннего фосфата очередного нуклеозидтрифосфата З'-ОН-группой на конце растущей цепи. В-третьих, движущая сила синтеза - гидролиз пирофосфата.
Рис. 25.8. Механизм реакции элонгации цепи, катализируемой РНК-по- лимеразой
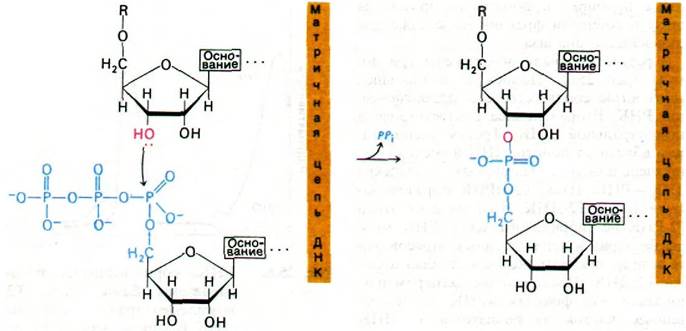
Однако по некоторым важным особенностям синтез РНК отличается от синтеза ДНК. Во-первых, РНК-полимеразе не нужна затравка. Во-вторых, ДНК-матрица при синтезе РНК полностью сохраняется, тогда как при синтезе ДНК она сохраняется лишь наполовину. В-третьих, РНК-полимераза, насколько известно, не обладает никакими нуклеазными активностями.
Все три типа клеточной РНК Е. coli - мРНК, тРНК и рРНК - синтезируются одной РНК-полимеразой в соответствии с инструкциями, заданными ДНК-матрицей. В клетках млекопитающих имеет место разделение труда между несколькими видами РНК-полимераз. Кроме того, необходимо отметить, что некоторые вирусы кодируют РНК-синтезирующие ферменты, совершенно отличные от ферментов клетки-хозяина. Таковы, например, РНК-полимераза, кодируемая ДНК-содержащим фагом Т7, и РНК-репликаза, кодируемая РНК-содержащим фагом Qß. Репликаза фага Qß является РНК-зависимой РНК- полимеразой, так как она синтезирует РНК не по ДНК-матрице, а по РНК (гл. 30). Клеточные же ферменты, синтезирующие РНК, наоборот, представляют собой ДНК-зависимые РНК-полимеразы.