БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 29. ЭУКАРИОТИЧЕСКИЕ ХРОМОСОМЫ И ВЫРАЖЕНИЕ ГЕНОВ У ЭУКАРИОТ
29.6. Нуклеосома - первый уровень конденсации ДНК
Накручивание ДНК на нуклеосомную сердцевину обеспечивает конденсацию ДНК, так как уменьшает ее линейные размеры. Участок ДНК длиной 200 пар оснований имеет в растворе длину 680 А. В нуклеосоме это количество ДНК укладывается в частицу диаметром 100 А. Таким образом, плотность упаковки (степень конденсации) нуклеосомы составляет примерно 7. Сравнима ли эта величина со степенью конденсации ДНК в хромосоме? Метафазные хромосомы человека, которые находятся в сильно конденсированном состоянии, содержат 5,3 • 109 пар оснований, что соответствует контурной длине ДНК 180 см. Эта ДНК упакована в 46 цилиндрических структур, общая длина которых составляет 200 мкм. Таким образом, плотность упаковки ДНК в метафазных хромосомах равна приблизительно 104. В интерфазных ядрах, где хроматин находится в более дисперсном состоянии, плотность упаковки для ДНК составляет примерно 102-103. Очевидно, нуклеосома представляет собой лишь первый уровень конденсации ДНК.
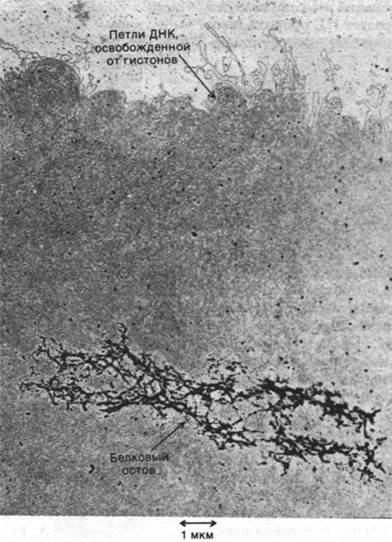
Каков следующий уровень организации ДНК? Одна из возможностей состоит в том, что сами нуклеосомы укладываются в спираль. Например, предложена соленоидная модель хроматина диаметром 360 А с плотностью упаковки около 40 (рис. 29.9). Складывание таких соленоидов в петли может дать дополнительную конденсацию. По всей вероятности, в стабилизации структуры хромосомы высших порядков участвуют различные негистоновые белки. Например, в лишенных гистонов метафазных хромосомах выявляется центральный белковый остов, окруженный множеством очень длинных петель ДНК (рис. 29.10). По- видимому, эти петли ДНК закреплены на остове. Такая организация может иметь важное значение для передвижения хромосом в митозе и мейозе.
Рис. 29.9. Предполагаемая соленоидная модель хроматина. На один оборот спирали приходится шесть нуклеосом (показаны синим цветом). Двойная спираль ДНК (красная) намотана на каждую нуклеосому
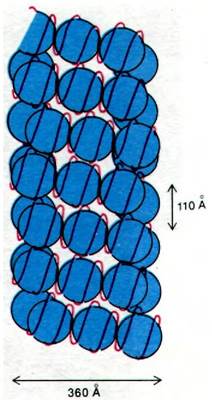
Рис. 29.10. Электронная микрофотография ДНК, с которой удалены гистоны. ДНК прикрепляется к центральному белковому остову. Гистоны удалили из метафазных хромосом клеток HeLa, обработав их полианионами
29.7. Репликация эукариотической ДНК начинается во многих местах и идет в двух направлениях
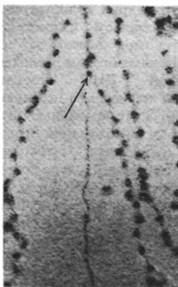
Подобно другим молекулам ДНК, эукариотическая ДНК реплицируется полуконсервативно. Кроме того, реплицированная ДНК полуконсервативно распределяется между сестринскими хроматидами метафазных хромосом (рис. 29.11). Методом электронной микроскопии было показано, что репликация эукариотической ДНК начинается во многих точках и идет в двух направлениях. Использование многих точек инициации необходимо для быстрой репликации, так как эукариотические ДНК имеют огромную длину. Например, самая длинная хромосома дрозофилы содержит ДНК длиной 2,1 см, или 62000 kb. Репликационная вилка ДНК дрозофилы движется со скоростью 2,6 kb/мин (сравните со скоростью движения вилки у Е. coli 16 kb/мин). Если бы в самой большой хромосоме дрозофилы существовала только одна точка начала репликации, то ее удвоение заняло бы более 16 сут. На самом деле время репликации составляет менее 3 мин. Это достигается кооперативным действием более чем 6000 репликационных вилок на одну молекулу ДНК. Молекула ДНК из ядра дрозофилы, находящегося на стадии дробления, представляет собой набор следующих один за другим участков репликации, или «глаз» (рис. 29.12). Активирование каждой точки инициации порождает две расходящиеся репликационные вилки. Расширяющиеся в обоих направлениях «глаза» сливаются и образуют две дочерние молекулы ДНК (рис. 29.13). «Глаз» внутри другого «глаза» никогда не наблюдался. Из этого следует что начало репликации не может снова реактивироваться, пока не закончится репликация всей молекулы ДНК.
Рис. 29.11. Флуоресцентная микрофотография метафазных хромосом мужчины. ДНК в клетках дважды реплицировалась в среде, содержащей аналог тимидина 5-бромдезоксиуридин (BrdU). Хромосомы сначала окрасили хинакрином (А), отмыли и снова окрасили бис- бензимидазольным красителем Хехст 33258 (Б). BrdU тушит флуоресценцию второго красителя, поэтому сестринские хроматиды, содержащие ДНК, в которой только одна полинуклеотидная цепь замещена BrdU, флуоресцирует ярче, чем ДНК с BrdU в обеих цепях
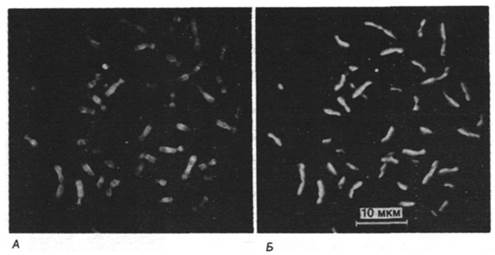
Рис. 29.12. Электронная микрофотография реплицирующейся хромосомной ДНК из дробящегося ядра эмбриона дрозофилы. «Глаза» - только что реплицированные участки
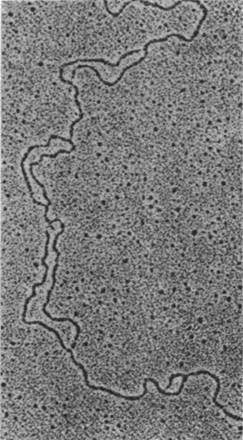
Рис. 29.13. Схематическое изображение репликации эукариотической хромосомы. Родительская ДНК показана синим цветом, новообразованная ДНК- красным
В эукариотических клетках имеются ДНК-полимеразы трех типов (табл. 29.3). α-Полимераза играет основную роль в репликации хромосомы, а β-фермент участвует в репарации ДНК. Когда покоящиеся клетки начинают быстро делиться, количество α-полимеразы возрастает более чем в 10 раз. γ-Полимераза отвечает за репликацию митохондриальной ДНК. Как и прокариотические ДНК-полимеразы, эти ферменты используют в качестве активированных предшественников дезоксирибонуклеозидтрифосфаты и осуществляют элонгацию затравки по матрице в направлении 5' → 3'. Но в отличие от прокариот эукариотические ДНК-полимеразы не обладают нуклеазной активностью. Скорее всего функцию исправления ошибок выполняют нуклеазы, ассоциированные с этими полимеразами в мультиферментных комплексах.
Таблица 29.3. Эукариотические ДНК-полимеразы

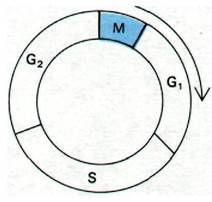
Рис. 29.14. Фазы клеточного цикла эукариотической клетки: митоз (М); фаза покоя 1, перед синтезом ДНК (G1); период синтеза ДНК (S); фаза покоя 2, после синтеза ДНК (G2). Продолжительность фаз зависит от типа клеток и условий культивирования. Митоз - обычно самая короткая фаза
29.8. Новые гистоны образуют новые нуклеосомы на отстающей дочерней цепи ДНК
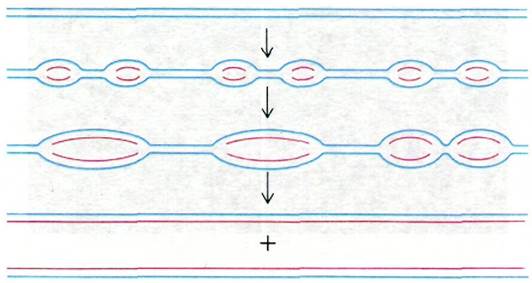
В репликационных вилках синтез ДНК идет в направлении 5' → 3' по одной дочерней цепи и в направлении 3' → 5' по другой. Как и при репликации прокариотической ДНК (разд. 24.19), это происходит следующим образом: одна цепь, ведущая, синтезируется непрерывно, а другая, отстающая, прерывисто. Как распределяются старые и новые гистоны? Чтобы решить этот вопрос, синтез ДНК проводили в присутствии ингибитора белкового синтеза циклогексимида. В этих условиях синтез ДНК прекращается в течение примерно 15 мин. При действии ДНКазы I половина новообразованной ДНК полностью распадается, а другая половина расщепляется на фрагменты длиной 200 пар оснований. Этот эксперимент, а также данные, полученные с помощью метки тяжелыми изотопами, показали, что родительские гистоны ассоциированы только с одной из двухспиральных дочерних ДНК, тогда как другая остается голой из-за отсутствия новых гистонов. Эта интерпретация находит прямое подтверждение на электронных микрофотографиях, где видно, что в области репликационной вилки одна из дочерних нитей покрыта бусинами, а другая - нет (рис. 29.15). Иными словами, родительские гистоны распределяются во время репликации консервативно1. Такое поведение гистонов показывает, что гистоны не отделяются от ДНК во время репликации. Старые гистоны остаются на двухцепочечной ДНК, содержащей ведущую цепь, тогда как новые гистоны садятся на ДНК, содержащую отстающую цепь. Причина этого различия между дочерними молекулами ДНК заключается, очевидно, в том, что гистоны гораздо прочнее связываются с двухцепочечной ДНК, чем с одноцепочечной. Старые гистоны, по-видимому, не удерживаются на отстающей цепи по той причине, что до соединения фрагментов Оказаки она содержит одноцепочечные участки.
Рис. 29.15. Асимметричная репликационная вилка, возникшая в результате синтеза ДНК в присутствии циклогексимида, блокирующего образование новых гистонов. Одна из дочерних молекул ДНК покрыта бусинами, другая остается голой. Это показывает, что родительские гистоны связаны только с одной из дочерних молекул
1 Недавно этот вывод подвергался серьезной критике в связи с тем, что по некоторым данным циклогексимид нарушает нормальное распределение гистонов во время репликации— Прим. перев.