БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
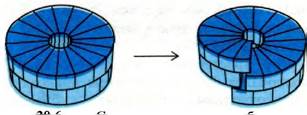
30.3. При сборке вирусной частицы ВТМ белковые диски присоединяются к петле РНК
A priori простейший механизм сборки ВТМ заключался бы в ступенчатом присоединении отдельных белковых субъединиц к РНК. Трудность, однако, состоит в этом случае в том, что скорость нуклеации будет чрезвычайно мала. Прежде чем комплекс замкнется сам на себя, образовав один оборот спирали, и таким образом стабилизируется, к гибкой молекуле РНК должно присоединиться примерно 17 субъединиц оболочки. Эта проблема может быть решена присоединением к РНК сразу целого комплекса из многих субъединиц. Действительно, белок оболочки легко образует двухслойный диск из 34 субъединиц. Каждый слой диска представляет собой кольцо из 17 субъединиц; примерно столько субъединиц (16 1/3) приходится на один оборот спирали ВТМ. Круг и его сотрудники исследовали трехмерную структуру диска (рис. 30.5) и показали, что он представляет собой главный промежуточный продукт при сборке ВТМ. Важнейшее свойство диска состоит в том, что его субъединицы скользят относительно друг друга и образуют спираль их двух оборотов, так называемую «запорную шайбу» (рис. 30.6).
Рис. 30.5. Карта электронной плотности белкового диска ВТМ. Показан слой толщиной 6 А

Рис. 30.6. Схема превращения белкового диска ВТМ в спиральную форму «запорной шайбы»
Диск гораздо быстрее взаимодействует с РНК ВТМ, чем с чужеродной ДНК. Следовательно, РНК ВТМ, по-видимому, содержит последовательность оснований, которую специфически узнает диск, и которая инициирует сборку. Эта область инициации была выделена следующим образом. К РНК ВТМ добавляли несколько дисков, чтобы покрыть область инициации, и затем расщепляли остальную РНК нуклеазой. Защищенный фрагмент содержит примерно 65 нуклеотидов, которые прочно и специфично связываются с дисками. Последовательность оснований в этой области инициации дает все основания думать, что этот фрагмент РНК образует структуру шпильки со стеблем, состоящим из спаренных оснований, и петлей (рис. 30.7). Самое интересное, что, петля содержит в каждом третьем положении G. Такое расположение оснований в триплетах соответствует стехиометрии субьединиц оболочки вируса - одна субъединица на три нуклеотида. Следовательно, по всей вероятности, петля связывается с первым диском, и с этого начинается сборка вирусной частицы.
Рис. 30.7. Участок РНК ВТМ, обеспечивающий инициирование сборки вирусной частицы ВТМ

Неожиданно оказалось, что, петля инициации расположена далеко от обоих концов РНК. Точка начала сборки расположена на расстоянии 5300 нуклеотидов от 5'-конца и примерно 1000 нуклеотидов от 3'-конца РНК. Другой неожиданностью было то, что оба конца РНК выходят с одной и той же стороны растущей частицы ВТМ (рис. 30.8). Длина 3'-конца на протяжении процесса сборки остается более или менее постоянной, а 5'-конец по мере удлинения вирусной частицы становится короче.
Рис. 30.8. Электронная микрофотография частично реконструированных частиц ВТМ. Видны два хвоста РНК, отходящие от каждого растущего вириона

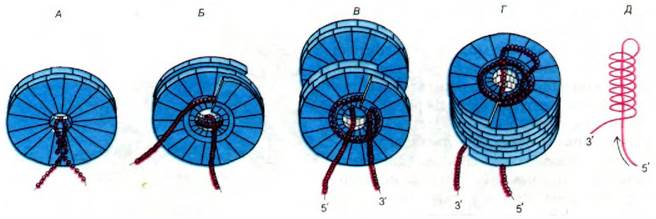
Наиболее вероятная модель образования частиц проиллюстрирована на рис. 30.9. Сборка начинается с проникновения петли инициации в центральное отверстие двухслойного белкового диска. Петля связывается с первым оборотом диска, и прилегающий стебель, состоящий из спаренных оснований, раскрывается. Это взаимодействие переводит диск в форму спиральной запорной шайбы, и диск захватывает РНК. Так начинается построение спирали вируса. Затем к новообразованной петле РНК, торчащей из центрального отверстия, присоединяется еще один диск. Благодаря протаскиванию 5'-конца через центральное отверстие растущей вирусной частицы при добавлении каждого следующего диска образуется новая петля. Наконец, происходит одевание 3'-конца каким-то пока непонятным способом.
Рис. 30.9. Схема сборки ВТМ. А - область инициации в РНК образует петлю и проходит в центральное отверстие белкового диска. Б - диск переходит в спиральную форму «запорной шайбы». В - к тому концу РНК, где расположена петля, присоединяются новые диски. Г - одна из концов РНК все время протаскивается через центральное отверстие и взаимодействует с новыми дисками. Д - схематическое изображение молекулы РНК в частично собранном вирусе. Направление движения РНК, обозначено стрелкой
Помимо того что двухслойный диск обеспечивает быструю нуклеацию, он существенно увеличивает специфичность образования оболочки. Диск может связываться со многими нуклеотидами, тогда как одна субъединица - только с тремя. Следовательно, диск обладает гораздо более высокой избирательностью по отношению к РНК ВТМ по сравнению с мРНК клетки-хозяина, чем одна субъединица. Еще одно важное свойство дисков состоит в том, что в физиологических условиях они не образуют спиралей без РНК. В этом отношении важнейшую роль играют две карбоксильные группы в каждой субъединице. При нейтральном значении рН в спиральной форме ионизированы обе карбоксильные группы, а в диске - только одна. Электростатическое отталкивание между близко расположенными карбоксилат-ионами в спиральной форме благоприятствует образованию диска. Связывание РНК со спиральной формой сопровождается достаточным изменением свободной энергии, чтобы преодолеть электростатическое отталкивание карбоксилатных ионов. Итак, карбоксилат- ионы - негативный регулятор, препятствующий образованию спирали без РНК.
30.4. Заражение фагом Т4 полностью перестраивает синтез макромолекул в клетке E. coli
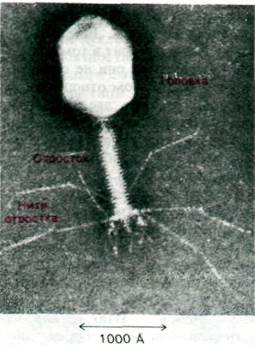
Бактериофаг Т4 - гораздо более сложный вирус, чем ВТМ. Его двухспиральная ДНК содержит примерно 165 генов по сравнению с 6 генами ВТМ. Однако структура, размножение и процесс сборки фага Т4 изучены довольно хорошо, так как он подвергался интенсивному генетическому и биохимическому анализу. Вирион Т4 состоит из головки. отростка и шести нитей (фибрилл) отростка (рис. 30.10). Его молекула ДНК плотно упакована внутри икосаэдрической белковой оболочки и образует головку вируса. Отросток состоит из двух соосных трубок, соединенных с головкой короткой шейкой. В отростке сократительный чехол окружает центральный стержень, через который ДНК вводится в бактерию-хозяина. Отросток несет на конце базальную пластинку с шестью короткими зубцами, от которой отходит шесть длинных тонких нитей.
Рис. 30.10. Электронная микрофотография фага Т4
Концы нитей отростка связываются с определенными участками на клетке E. coli. В результате АТР-зависимого сокращения чехол подтягивает головку фага к базальной пластинке и нитям отростка, и в результате центральный стержень проникает через клеточную стенку, но не через мембрану клетки. Затем обнаженная фаговая ДНК проникает через клеточную мембрану. По истечении несколько минут все реакции синтеза клеточных ДНК, РНК и белка останавливаются и начинается синтез вирусных макромолекул. Другими словами, заразивший клетку вирус овладевает синтетическими механизмами бактериальной клетки и замещает ее гены своими.
Таблица 30.2. Гены фага Т4

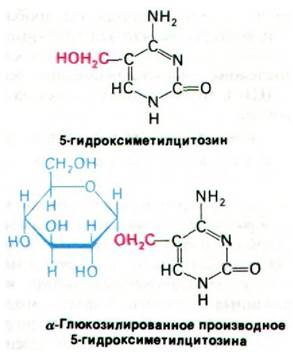
В ДНК фага Т4 имеется три группы генов, которые транскрибируются на различных стадиях заражения: предранние, ранние и поздние. Предранние и ранние гены транскрибируются и транслируются до того, как синтезируется ДНК фага Т4. Некоторые белки, кодируемые этими генами, обеспечивают выключение синтеза клеточных макромолекул. Вскоре после заражения ДНК клетки- хозяина распадается под действием дезоксирибонуклеазы, кодируемой одним из ранних генов фага Т4. ДНК самого фага Т4 не гидролизуется под действием этого фермента, поскольку в ней нет кластеров (сгруппированных остатков) цитозина. В ДНК фага Т4 вместо цитозина находится гидроксиметилцитозин(ГМЦ). К тому же остатки ГМЦ в ДНК Т4 глюкозилированы.
Эти производные цитозина включаются в ДНК бактериофага Т4 благодаря действию нескольких фагоспепифических ферментов, синтезирующихся на ранней стадии заражения. Один из них гидролизует dСТР с образованием dСМР, чтобы воспрепятствовать включению dСТР в ДНК фага Т4. Затем второй фермент вводит в dСМР гидроксиметильную группу, и образуется 5-гидроксиметилцитидилат. Третий фермент превращает 5-гидроксиметилцитиди- лат в трифосфат, который служит субстратом для ДНК-полимераз. Наконец, четвертый фермент гликозилирует некоторые из содержащихся в ДНК остатки гидроксиметилцитозина.
Синтез поздних белков сопряжен с репликацией ДНК фага Т4. На этом этапе образуются белки капсида и лизоцим. Когда сборка вирионов потомства завершена, лизоцим гидролизует клеточную стенку бактерии и разрушает ее. Примерно через 20 мин после заражения возникает около двухсот новых вирусных частиц.