БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
30.8. Гибкость белка оболочки ВККТ позволяет ему образовывать икосаэдрический капсид
Вирус кустистостой карликовости томатов (ВККТ) - сферический вирус, иллюстрирующий другой принцип организации вируса (рис. 30.15). ВККТ содержит одну молекулу РНК длиной 4800 нуклеотидов, окруженную оболочкой из 180 идентичных белковых субъединиц массой 41 кДа. Как уложены эти субъединицы оболочки? Максимально возможная симметрия изотермической оболочки достигается в случае икосаэдра и имеет порядок 60 (рис. 30.16, А). Другими словами, не более 60 идентичных субъединиц могут быть упакованы в сферическую оболочку с соблюдением абсолютной симметрии. Но ВККТ и ряд других сферических вирусов содержат 180 идентичных субъединиц. Биологическое преимущество построения оболочки из 180 вместо 60 субъединиц такого же размера состоит в том, что в более крупный вирион можно упаковать больше нуклеиновой кислоты. Это достигается не нарушением симметрии, а некоторым ее ослаблением (рис. 30.16, Б).
Рис. 30.15. Электронная микрофотография вируса кустистой карликовости томата
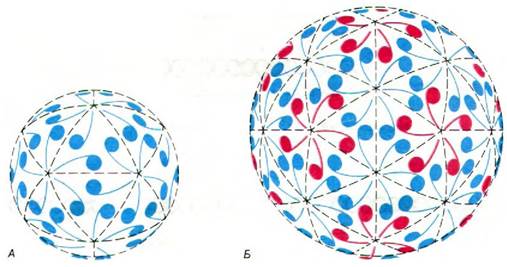
Рис. 30.16. Икосаэдрическая поверхностная решетка, демонстрирующая упаковку 60 совершенно одинаковых субъединиц (А) и 180 квазиэквивалентных субъединиц (Б). Обратите внимание, что все контакты типа «хвост к хвосту» на рис. А образуются кольцевыми группами по пять субъединиц, а на рис. Б некоторые такие контакты образуются группами по пять, а другие — по шесть субъединиц
Рентгеноструктурный анализ структуры ВККТ высокого разрешения, проведенный Стивеном Харрисоном (Stephen Harrison), показал, что химически идентичные субъединицы его оболочки можно отнести к трем группам (А, В и С), каждая из которых состоит из 60 белков, подчиняющихся строгой икосаэдрической симметрии. В то же время молекулы из разных групп расположены по отношению друг к другу несколько по-разному; поэтому их называют квазиэквивалентными.
Как объяснить квазиэквивалентность с физической точки зрения? Рентгеноструктурный анализ показал, что каждая субъединица состоит из S-домена, образующего часть поверхности оболочки; Р-домена, выступающего наружу; N-концевого участка, направленного внутрь. Р- и S-домены всех субъединиц имеют примерно одинаковое строение. В то же время угол между Р- и S- доменами в субъединицах группы С сильно отличается от угла в субъединицах А и В. Ри S-домены соединены шарниром, обеспечивающим поворот на угол до 20°. Еще одно отличие состоит в том, что N-концевая часть в субъединицах группы С упорядочена, а в субъединицах А и В находится в беспорядочной конформации. Благодаря такой гибкости структурной организации субъединицы, принадлежащей к различным группам, могут взаимодействовать друг с другом почти идентичным образом, что необходимо для самосборки. С другой стороны, РНК внутри частицы, по-видимому, не находится в какой-нибудь определенной конформации. Гибкие N-концевые участки субъединиц оболочки проникают внутрь и взаимодействуют с РНК.
30.9. Бактериальные рестрикционные эндонуклеазы расщепляют чужеродные молекулы ДНК
Мы уже видели, что фаг Т4 обладает ферментативной системой для избирательного расщепления ДНК клетки-хозяина. Подобно этому, у бактерий имеются ферменты, называемые рестриктирующими эндонуклеазами, которые расщепляют чужеродные молекулы ДНК. Эти ферменты были открыты в результате наблюдения, что фаги, выращенные на одном штамме бактерий (например, E. coli В), плохо растут на другом штамме (E. coli К) и наоборот. Такое явление, когда фаг плохо растет на штамме, отличном от того, на котором он был выращен, было названо рестрикцией. Однако небольшая часть фага (примерно 10-5) избегает рестрикции и в дальнейшем хорошо растет на новом хозяине. При этом фаги теряют способность расти на старом хозяине.
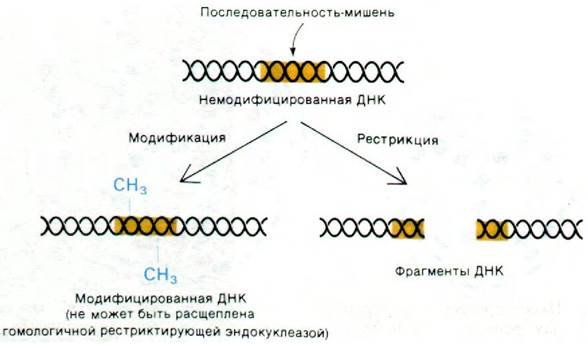
Все эти данные показали, что специфическая модификация в клетках-хозяевах защищает фаг от рестрикции. Затем Вернер Арбер (Werner Arber) продемонстрировал, что специфическая модификация клеткой-хозяином на самом деле воздействует на фаговую ДНК и что рестрикция обусловлена деградацией ДНК фага. ДНК клетки-хозяина и другие молекулы ДНК, содержащиеся в клетках-хозяевах, метилированы по определенным участкам. Те же участки узнает и рестриктирующая эндонуклеаза, которая расщепляет только неметилированные последовательности. Таким образом, метилирование определенного основания в последовательности-мишени (участке узнавания) препятствует гидролизу ферментом рестрикции (рис. 30.17). Момент, в который происходит модификация, имеет принципиально важное значение: бактериальная ДНК не подвергается расщеплению, потому что она метилируется раньше. Только что реплицированная бактериальная хромосома, метилированная только по одной - родительской - цепи, устойчива к действию фермента рестрикции. Такая наполовину модифицированная ДНК становится полностью метилированной до начала следующего цикла репликации.
Рис. 30.17. Метилирование участков узнавания (последовательностей- мишеней) защищает их от расщепления рестриктирующей эндонуклеазой
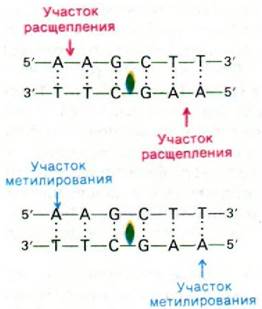
У бактерий обнаружено два типа систем рестрикция-модификация. В системах типа I активность метилазы и нукдеазы ассоциирована с крупным комплексом, состоящим из нескольких субъединиц. Например, такие комплексы у E. coli К и В состоят из полипептидных цепей трех типов. α-Цепь обладает эндонуклеазной активностью, ß-цепь - метилазной активностью, а γ-цепь несет участок узнавания ДНК. Ферменты типа I нуждаются в S-аденозилметионине и в АТР для проявления как нуклеазной, так и метилазной активности. Ферменты типа I расщепляют не модифицированную ДНК в случайном месте на расстоянии 1900 пар оснований или более в 5'-сторону от участка узнавания1 и одновременно гидролизуют АТР. В системах типа II, наоборот, метилазы и нуклеазы разделены. S-аденозилметионин служит донором метальной группы в реакции модификации, но не участвует в расщеплении ДНК. Еще одно отличие состоит в том, что нуклеазы и метилазы типа II не нуждаются в АТР. Самое удивительное, что места расщепления нуклеазами типа II весьма специфичны. Как уже обсуждалось в одной из предыдущих глав (разд. 24.27), многие из этих ферментов узнают определенную последовательность из 4-6 пар оснований и гидролизуют в каждой цепи единственную строго определенную фосфодиэфирную связь в этой области. Отличительная особенность этих участков расщепления состоит в том, что они симметричны относительно оси вращения второго порядка (рис. 30.18). Ферменты рестрикции - незаменимый инструмент в исследовании структуры ДНК (разд. 24.27) и создании новых молекул ДНК (разд. 31.9).
1 Арбер изучал систему рестрикции—модификации типа I на содержащем одноцепопечную ДНК фаге fd, поэтому он мог указать положение участка расщепления относительно участка узнавания так, как это сделано в тексте. Прим. перев.
Рис. 30.18. Специфичность рестриктирующей эндонуклеазы Hind III из Hemophilus influenza и соответствующей метилазы. Зеленым цветом показана ось симметрии второго порядка
30.10. Стратегия репликации РНК-содержащих вирусов
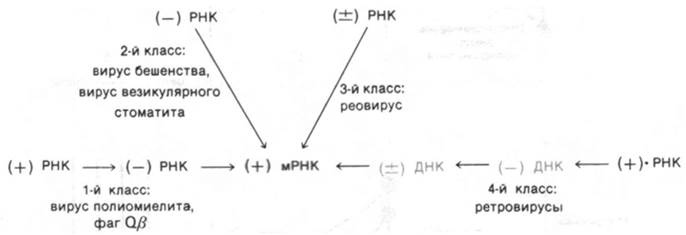
Репликация РНК-содержащих вирусов представляет собой особую проблему, так как в незараженных клетках-хозяевах нет ферментов для синтеза РНК по РНК-матрице. Следовательно, РНК-содержащие вирусы должны нести генетическую информацию для синтеза РНК-зависимой РНК- полимеразы (которую называют также РНК-репликазой или РНК-синтетазой) или для РНК-зависимой ДНК-полимеразы (называемой также обратной транскриптазой). РНК-содержащие вирусы удобно классифицировать в зависимости от РНК, содержащейся в вирионе, и мРНК. По определению мРНК представляет собой (+) РНК, а комплементарная ей цепь - (-) РНК. Известны четыре пути репликации и транскрипции РНК-содержащих вирусов (рис. 30.19). Вирусы 1-го класса (например, вирус полиомиелита) - вирусы, содержащие положительную цепь РНК. Они синтезируют (-) РНК, которая используется в качестве матрицы для образования (+) мРНК. Вирусы 2-го класса (например, вирусы бешенства) содержат отрицательную цепь РНК, у них (-) РНК вириона служит матрицей для синтеза (+) мРНК. Вирусы 3-го класса (например, реовирусы) содержат двухцепочечную РНК; (±) РНК вириона направляет асимметрический синтез (+) мРНК. Наиболее необычны вирусы 4-го класса - ретровирусы (например, вирус саркомы Рауса). У них выражение генетической информации, содержащейся в (+) РНК вириона, опосредовано образованием ДНК, которая служит матрицей для синтеза (+) РНК. Таким образом, поток генетической информации ретровирусов направлен от РНК к ДНК и затем обратно к РНК.
Рис. 30.19. Способы экспрессии генов РНК-содержащих вирусов
Пикорнавирусы - группа мелких (pica) РНК-содержащих (рна - от айгл. RNA) вирусов. Одна одноцепочечная молекула (+) РНК окружена икосаэдрической белковой оболочкой диаметром 270 А. К пикорнавирусам относятся вирус полиомиелита, риновирус (вызывающий обычную простуду) и вирус ящура крупного рогатого скота.