БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
30.15. Дарвиновская эволюция фаговой РНК вне клетки
Очистка РНК фага Qβ и Qβ-репликазы от примесей нуклеаз позволила Солу Спигелману (Sol Spigelman) изучать эволюционные события вне живой клетки. Один из вопросов был сформулирован следующим образом: что произойдет с молекулами РНК, если единственное предъявляемое к ним требование - это как можно быстрее размножаться? Молекулы РНК и репликазы фага Qβ и рибонуклеозидтрифосфаты инкубировали в течение 20 мин. Такое время инкубации способствует отбору мутантных молекул РНК, которые быстро реплицируются. Образец этой инкубационной смеси переносили и разводили в свежей порции стандартной реакционной смеси, содержащей Qβ-репликазу и рибонуклеозидтрифосфаты. Проводили 75 переносов и затем анализировали образовавшиеся РНК. Продолжительность инкубации постепенно снижали, так как молекулы РНК на протяжении эксперимента реплицировались все быстрее. Самым удивительным было то, что после 75 переносов («поколений») длина молекул РНК составляла всего 12% длины исходной РНК фага Qβ. Нуклеотиды, ненужные для репликации, были утрачены, так как укороченные молекулы реплицировались быстрее. Основное ограничение, которое накладывали условия этого эксперимента, сохранение в мутантных молекулах инициирующей последовательности, которую узнает Qβ-репликаза.
30.16. Лизогенные фаги могут включать свою ДНК в состав ДНК клетки-хозяина
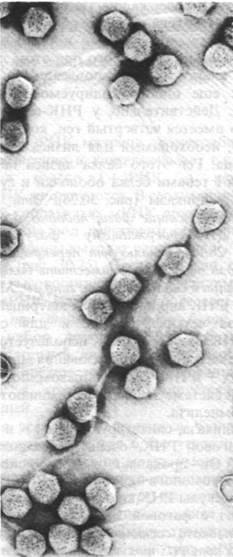
У некоторых бактериофагов существует два возможных пути, по которым может пойти их дальнейшее развитие после заражения клетки-хозяина: они могут размножаться и лизировать зараженную клетку (литический путь развития) или же их ДНК может включиться в ДНК зараженной клетки, не проявляя способности к размножению и лизису (лизогенный путь развития). Вирусы, которые не всегда убивают клетку-хозяина, называются умеренными. Лучше всего из умеренных вирусов изучен фаг X (рис. 30.27); мы уже говорили о нем раньше в связи с регуляцией транскрипции (разд. 28.11). Напомним, что репрессор фага X связывается с двумя группами операторных участков OL и OR и что он регулирует свой собственный синтез.
Рис. 30.27. Электронная микрофотография фага ![]()
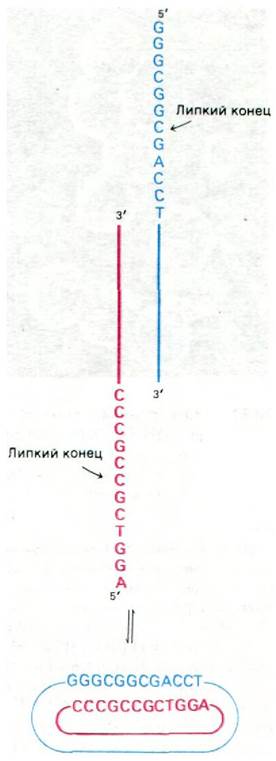
ДНК вириона - линейная двухцепочечная молекула длиной 48 кb. 5'-конец каждой ее цепи представляет собой одноцепочечную последовательность из 12 нуклеотидов. Эти последовательности называются липкими концами, так как они взаимно комплементарны и могут спариваться друг с другом. На самом деле они соединяются сразу после заражения. В результате 5'-фосфат каждой цепи оказывается рядом со своим собственным З'-гидроксильным концом. ДНК-лигаза клетки-хозяина заделывает разрывы, и в результате образуется кольцевая молекула ДНК фага ![]() (рис. 30.28).
(рис. 30.28).
Рис. 30.28. Превращение линейной ДНК фага ![]() в кольцевую форму
в кольцевую форму
Репликация этой кольцевой молекулы ДНК фага X происходит путем взаимодействия белков, кодируемых фагом ![]() , с репликационными механизмами клетки-хозяина. В другом случае кольцевая ДНК фага X может включиться в бактериальную хромосому с помощью одного акта реципрокной рекомбинации между специфическими участками ДНК фага
, с репликационными механизмами клетки-хозяина. В другом случае кольцевая ДНК фага X может включиться в бактериальную хромосому с помощью одного акта реципрокной рекомбинации между специфическими участками ДНК фага ![]() и E. coli длиной по 15 пар основании. Сайт (участок) прикрепления фага
и E. coli длиной по 15 пар основании. Сайт (участок) прикрепления фага ![]() в ДНК E. coli обозначается attX и располагается между генами галактозного и биотинового оперонов galE и biоА.Последовательность оснований attX можно символически обозначать В-В' (от англ. bacterial - бактериальный). Специфический сайт прикрепления в фаге X называется att и локализован рядом с генами int (от англ. integrate - интегрировать) и xis (от англ. excise - вырезать). Последовательность оснований att обозначается Р-Р' (от англ. phage - фаг). Белок int узнает последовательность Р-Р' в фаговой ДНК и последовательность В-В' в ДНК E. coli. Затем происходит взаимный перенос: Р соединяется с В', а В - с P'. Эта схема (рис. 30.29 и 30.30) была первоначально предложена Алланом Кэмпбеллом (Allan Campbell) на основании генетических данных.
в ДНК E. coli обозначается attX и располагается между генами галактозного и биотинового оперонов galE и biоА.Последовательность оснований attX можно символически обозначать В-В' (от англ. bacterial - бактериальный). Специфический сайт прикрепления в фаге X называется att и локализован рядом с генами int (от англ. integrate - интегрировать) и xis (от англ. excise - вырезать). Последовательность оснований att обозначается Р-Р' (от англ. phage - фаг). Белок int узнает последовательность Р-Р' в фаговой ДНК и последовательность В-В' в ДНК E. coli. Затем происходит взаимный перенос: Р соединяется с В', а В - с P'. Эта схема (рис. 30.29 и 30.30) была первоначально предложена Алланом Кэмпбеллом (Allan Campbell) на основании генетических данных.
Рис. 30.29. схема реципрокной рекомбинации ДНК фага ![]() и ДНК Е. coli
и ДНК Е. coli
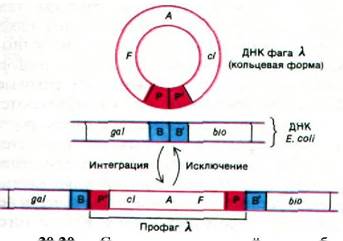
Рис. 30.30. Интеграция и исключение ДНК фага X. Это неполная схема, так как некоторые факторы, участвующие в этих процессах, пока не идентифицированы
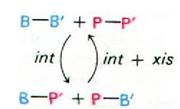
Теперь ДНК фага X составляет часть молекулы ДНК E. coli. Эта форма называется профагом, а клетка E. coli, содержащая профаг, лизогенной бактерией. Профаг стабилен в отсутствие белка xis. Транскрипция гена xis блокируется репрессором фага X (разд. 28.11). Когда репрессия снимается. белки xis и int совместно катализируют разрыв последовательностей В—Р' и Р—В', и снова происходит взаимный перенос (рис. 30.30): Р соединяется с Р', а В с В'; при этом снова получаются кольцевая молекула ДНК фага X и нелизогенная хромосома E. coli. Главная особенность этой системы рекомбинации заключается в том. что белок int сам по себе не может узнавать две новые последовательности на концах профага (В-Р' и Р-В'), поэтому они устойчивы. Таким образом, внедрение фага происходит в присутствии одного белка int, тогда как вырезание профага - только в присутствии обоих белков - int и xis.
30.17. Ретровирусы и некоторые ДНК- содержащие вирусы могут вызывать рак у чувствительных клеток-хозяев
В 1911 г. Пейтон Раус (Peyton Rous) приготовил бесклеточный фильтрат из опухоли соединительной ткани, которая спонтанно возникла у курицы и ввел его нормальным цыплятам. Как ни странно, у реципиентов развились высокозлокачественные опухоли такого же типа, которые называются саркомами. Кроме того, Раус обнаружил, что опухолеродный фактор фильтрата, известный в настоящее время под названием вируса саркомы Рауса (RSV) или вируса саркомы птиц (ASV) (рис. 30.31), можно размножить путем последовательного пассирования в курах. Вирус саркомы птиц относится к группе РНК-содержащих опухолеродных вирусов (онкогенных РНК-содержащих вирусов). Эти вирусы содержат в составе виринов (+) РНК и размножаются с использованием двухспиральной ДНК-посредника. Поэтому их называют ретровирусы. Ретровирусы - единственные РНК-содержащие вирусы, способные вызывать рак. Кроме того, злокачественные опухоли может вызывать ряд ДНК-содержащих вирусов. Обезьяний вирус 40 (SV-40) и вирус полиомы принадлежат к группе паповавирусов (рис. 30.32); из всех онкогенных ДНК-содержащих вирусов они изучаются наиболее интенсивно. Особый интерес к этим РНК- и ДНК-содержащим вирусам привлекает то, что они содержат всего четыре-пять генов. В индукции рака участвует всего один или два вирусных гена, и потому исследователи питают надежду выяснить механизм их действия.
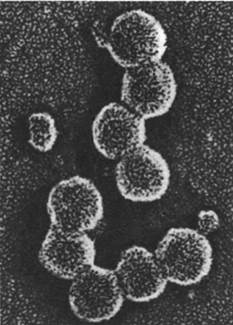
Рис. 30.31. Электронная микрофотография вируса саркомы птиц (вируса саркомы Рауса), относящегося к ретровирусам. Сильноокрашивающиеся вирионы расположены вблизи поверхности зараженной клетки цыпленка
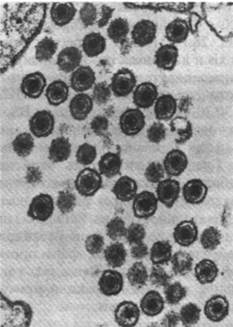
Рис. 30.32. Электронная микрофотография ДНК-содержащего опухолеродного вируса SV-40
Для изучения рака на молекулярном уровне были разработаны системы тканевых культур. При проникновении онкогенного вируса в подходящие животные клетки они становятся постоянно трансформированными, т.е. похожими на раковые. Трансформированные клетки отличаются от нормальных, особенностями своего роста и характером клеточных поверхностей (табл. 30.3). Самое разительное изменение состоит в том, что трансформированные клетки растут непрерывно и хаотически независимо от соседних клеток. Кроме того, трансформированные клетки содержат вирусоспецифическую ДНК, интегрированную с геномом клетки-хозяина. Этим объясняется тот факт, что трансформация является наследуемым изменением фенотипа. Культуры, полученные из трансформированных колоний, навсегда сохраняют аномальные свойства трансформированных клеток. Кроме того, некоторые трансформированные клетки, полученные из культуры ткани, при введении в достаточном количестве в подходящего хозяина образуют раковую опухоль.
Таблица 30.3. Изменения свойств клеток при трансформации ДНК- или РНК-содержащими опухолероднымн вирусами
Характер роста
При введении чувствительным животным образуют опухоли
Растут до гораздо более высокой плотности Рост становится не ориентированным в пространстве и клетки отделяются от поверхности, на которой растут
В среду выделяются активаторы протеаз, что повышает инвазивность клеток
Снижается потребность в факторах роста сыворотки
Свойства поверхности
Появляются новые вирусоспецифические антигены
Один из белков наружной поверхности клеток - фибронектин - исчезает
Падает содержание ганглиозидов
На поверхности клетки появляются антигены плода
Скорость транспорта питательных веществ увеличивается
Способность к агглютинации под действием растительных лектинов увеличивается
Признаки присутствия опухолеродного вируса
Имеются последовательности вирусной ДНК
Имеются вирусоспецифические мРНК
Выявляются вирусоспецифические антигены