БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 31. ПЕРЕСТРОЙКИ ГЕНОВ: РЕКОМБИНАЦИЯ, ТРАНСПОЗИЦИЯ И КЛОНИРОВАНИЕ
Тема настоящей главы - перестройка генов путем перемещения крупных участков ДНК. Прежде всего мы обсудим процесс генетической рекомбинации, при котором новая молекула ДНК возникает путем разрыва и воссоединения цепей ДНК. Вероятность генетической рекомбинации существенно увеличивается при наличии обширных участков гомологии между взаимодействующими молекулами ДНК. Были выделены промежуточные продукты рекомбинации, а сравнительно недавно был охарактеризован и фермент, катализирующий взаимный обмен цепей ДНК. Затем мы обсудим транспозицию - перемещение гена из одной хромосомы в другую или с одного места на другое в пределах одной хромосомы. В отличие от общей рекомбинации для транспозиции не нужны протяженные участки гомологии. У прокариот присутствие так называемых последовательностей-вставок, или IS-элементов (от англ. insertion sequences), сообщает подвижность неродственным фрагментам ДНК, обеспечивая их соединение. Рекомбинация и транспозиция сыграли важную роль в эволюции, так как они приводили к возникновению новых геномов. В конце настоящей главы мы рассмотрим конструирование новых комбинаций генов в пробирке и их выражение в клетках-хозяевах. Гены можно ковалентно соединить с ДНК плазмид и вирусов с помощью рестриктирующих эндонуклеаз и ДНК-лигазы. Такие рекомбинатные молекулы ДНК могут реплицироваться и экспрессироваться в подходящих клетках-хозяевах. Кроме того, мы обсудим важность клонирования генов и возможность его практического применения. Исследования рекомбинации и транспозиции и разработка методов клонирования идут исключительно быстрым темпом. В результате этих работ возникают новые плодотворные методы изучения геномов, которые позволяют глубже проникнуть в механизмы их эволюции и выражения.
31.1 В основе генетической рекомбинации лежат разрыв и воссоединение цепей ДНК
При генетической рекомбинации возникает молекула ДНК, последовательность которой происходит частично от одной родительской молекулы, а частично от другой. Исследования клеток Е. coli, зараженных смесью фагов Т4, меченных 32Р и бром- дезоксиурацилом, позволили получить представление о молекулярной природе рекомбинантных молекул. Плавучая плотность ДНК, меченной бромурацилом, значительно выше, чем плотность ДНК, меченной Р, так что родительские молекулы можно отделить друг от друга и от рекомбинантных молекул центрифугированием в градиенте плотности хлористого цезия (CsCl). Результаты опытов по центрифугированию показали, что после заражения смесью фагов рекомбинантные молекулы содержат и 32Р, и бромурацил. Структура гибридных молекул зависела от того, происходил ли во время их образования синтез ДНК. В отсутствие синтеза ДНК 32Р-ДНК в составе рекомбинантных молекул не была ковалентно соединена с меченной бромурацилом ДНК. При нагревании выше температуры плавления двухспиральных молекул гибрид диссоциировал на легкий и тяжелый компоненты. Фрагменты родительских молекул в этом гибриде удерживаются вместе в результате спаривания оснований; поэтому этот промежуточный продукт называется составным (рис. 31.3). Если же синтез ДНК происходил, одноценочечные пробелы в составном промежуточном продукте заполнялись ДНК-полимеразой I и концы соединялись ДНК-лигазой. Образовавшуюся рекомбинатную молекулу уже нельзя было разделить на легкий и тяжелый компоненты, так как куски ДНК были ковалентно соединены. Эти эксперименты показали, что одноцепочечные участки ДНК являются промежуточными продуктами генетической рекомбинации и что в этом процессе участвуют ферменты.
Рис. 31.1. Перенос генетической информации от одной клетки Е. coli к другой. На этой электронной микрофотографии видны две клетки Е. coli, соединенные при помощи пиля во время конъюгации. ДНК переносится через пиль из донорной клетки в акцепторную
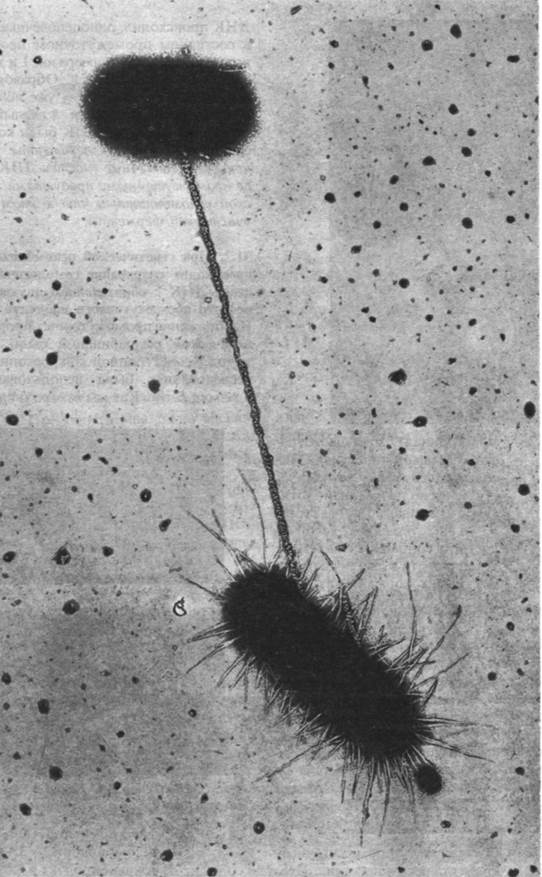
Рис. 31.2. Электронная микрофотография кольцевой молекулы ДНК, содержащей гены устойчивости к нескольким антибиотикам. Такие плазмиды (факторы R - от англ. resistance - устойчивость) сообщают клеткам устойчивость к различным веществам, которая может передаваться другим клеткам
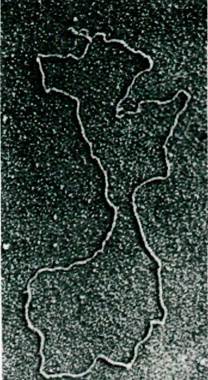
Рис. 31.3. Составной промежуточный продукт рекомбинации ДНК фага Т4 в зараженных бактериях. Меченная 32Р ДНК показана красным цветом, а ДНК, меченная бромурацилом -синим