БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 31. ПЕРЕСТРОЙКИ ГЕНОВ: РЕКОМБИНАЦИЯ, ТРАНСПОЗИЦИЯ И КЛОНИРОВАНИЕ
31.5. Фактор F позволяет бактериям передавать гены реципиентам путем конъюгации
Некоторые плазмиды обусловливают перенос бактериями генетического материала в другие бактерии путем образования непосредственных межклеточных контактов. Этот процесс, названный конъюгацией, открыли в 1946 г. Джошуа Ледерберг и Эдвард Татум (Joshua Lederberg, Edward Tatum). При конъюгации клеток Е. coli один партнер (мужского пола) служит донором генетического материала, другой (женского пола) - реципиентом. Бактерии мужского пола имеют на поверхности особые отростки, получившие название половых пилей, а женские клетки несут рецепторные участки, которые связывают пили. Пиль связывает между собой мужскую и женскую клетки (рис. 31.1). Затем он сокращается, что позволяет клеткам вступить в непосредственный контакт для передачи ДНК (рис. 31.10). Бактерия мужского пола содержит плазмиду, называемую фактором F (от англ. fertility - плодовитость), которая несет гены, детерминирующие образование половых пилей и других компонентов, участвующих в конъюгации. При конъюгации одна цепь плазмиды фактора Fразрывается в одном месте, и происходит расплетание двухцепочечной молекулы (рис. 31.11). 5'-конец разорванной цепи входит в реципиентную клетку, и на ней синтезируется комплементарная цепь. При этом образуется замкнутая кольцевая двухцепочечная молекула. Присутствие плазмиды фактора F в реципиентной клетке (первоначально F) превращает ее в мужскую клетку (F+). Мужская клетка может спонтанно терять свой фактор и ревертировать таким образом к генотипу F-.
Рис. 31.10. Электронная микрофотография двух клеток Е. coli во время конъюгации
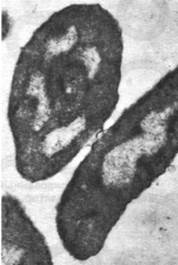
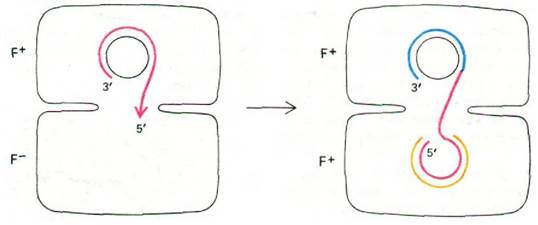
Рис. 31.11. Предполагаемый механизм переноса нити фактора R при конъюгации. В суперспирализованную молекулу фактора R в клетке F+-донора вносится одноцепочечный разрыв, и после этого он расплетается, Перенос одной цепи фактора R в клетку F--реципиента сопряжен с репликацией этой цепи донора. Затем в реципиентной клетке синтезируется комплементарная цепь
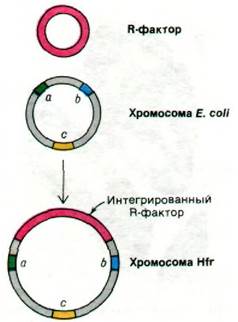
Плазмида фактор F может интегрировать с бактериальной хромосомой. Интеграция происходит путем кроссинговера с одним из множества мест в бактериальной хромосоме. Частота интеграции составляет примерно 10-5 в расчете на одну генерацию. Бактерии, несущие фактор F в своих хромосомах, называются клетками Hfr (от англ. high frequency ofrecombination - высокая частота рекомбинации). Клетки Hfr, как и клетки F+, участвуют в конъюгации в качестве доноров (рис. 31.12). Различие между ними состоит в том, что клетка Hfr передает всю бактериальную хромосому (включая интегрированный фактор F), тогда как клетка F+ передает реципиенту только фактор F. При скрещивании Hfr x F- вся хромосома переносится примерно за 90 мин. Порядок входа передаваемых генов в реципиентную клетку зависит от места, в котором произошла интеграция фактора F и от его полярности. Поэтому порядок генов в хромосоме донора можно легко установить, прерывая конъюгацию в различные моменты времени и определяя, какие маркеры успели перейти. Передаваемая хромосома донора может рекомбинировать с хромосомой реципиента. Частота рекомбинации выше всего для генов, которые вошли в реципиентную клетку первыми, так как они находятся там дольше всего. Таким образом, можно строить генетические карты, определяя время входа и частоту рекомбинации маркеров, передаваемых донором.
Рис. 31.12. Схема образования Нfг-клеток путем интеграции фактора F с хромосомой Е. coli
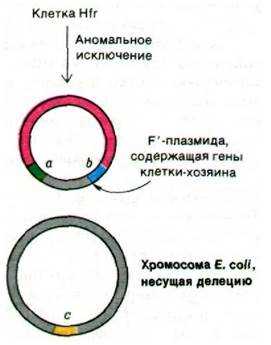
При исключении фактора F из хромосомы Hfr-клетки она переходит в состояние F+. Этот процесс, обратный интеграции фактора F, также происходит с частотой примерно 10-5 за одну генерацию. В небольшой части ревертантов кроссинговер происходит в сайте, отличном от сайта интеграции. В результате образуется плазмида, содержащая, помимо генов фактора F, хромосомные гены (рис. 31.13). Такая плазмида называется фактором F'; штрих обозначает, что в ней присутствуют хромосомные гены. Конъюгация клетки F' с клеткой F-приводит к переносу этих хромосомных генов из донора в реципиентную клетку, которая в результате становится диплоидной по этим генам.
Рис. 31.13. Аномальное исключение фактора F приводит к образованию плазмиды F', содержащей часть хромосомы Е. coli
Таким образом, у бактерий имеется механизм для переноса целых хромосом или нескольких генов из одной клетки в другую. Фактор F можно рассматривать как особый переносчик (вектор), возникший специально для обмена генетическим материалом. Интересно отметить, что лизогенизирующие фаги также могут участвовать в обмене генов клетки-хозяина. Например, ДНК фага ![]() может интегрировать между генами gal и bio хромосомы Е. coli (разд. 30.16). Исключение профага из хромосомы обычно происходит точно, но не всегда. Примерно в одном вирионе на 105 ДНК фага
может интегрировать между генами gal и bio хромосомы Е. coli (разд. 30.16). Исключение профага из хромосомы обычно происходит точно, но не всегда. Примерно в одном вирионе на 105 ДНК фага ![]() содержит оперон gal или ген bio. При заражении такими фагами, называемыми
содержит оперон gal или ген bio. При заражении такими фагами, называемыми ![]() gal и
gal и ![]() bio, эти гены вводятся в клетку Е. coli вместе с генами фага
bio, эти гены вводятся в клетку Е. coli вместе с генами фага ![]() . Другой родственный фаг, который называется ɸ80, интегрирует вблизи оперона trp и может переносить гены trp из одной зараженной клетки в другую. Бактериофаг μинтегрирует почти в любом месте хромосомы Е. coli и при исключении всегда захватывает кусок бактериальной хромосомы. Эти трансдуцирующие фаги, подобно фактору F, представляют собой подвижные генетические элементы, которые обеспечивают взаимообмен бактериальных генов. Не исключено, что трансдукция ускоряет эволюцию бактерий.
. Другой родственный фаг, который называется ɸ80, интегрирует вблизи оперона trp и может переносить гены trp из одной зараженной клетки в другую. Бактериофаг μинтегрирует почти в любом месте хромосомы Е. coli и при исключении всегда захватывает кусок бактериальной хромосомы. Эти трансдуцирующие фаги, подобно фактору F, представляют собой подвижные генетические элементы, которые обеспечивают взаимообмен бактериальных генов. Не исключено, что трансдукция ускоряет эволюцию бактерий.
31.6. Плазмиды факторы R придают бактериям устойчивость к антибиотикам
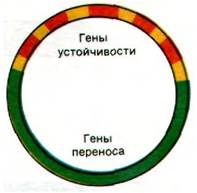
Поразительным примером необычно быстрой эволюции бактерий может служить эпидемия бактериальной дизентерии, протекавшая в 1955 г. Один из штаммов Shigella dysenteriaeприобрел устойчивость одновременно к хлорамфениколу, стрептомицину, сульфаниламидам и тетрациклину. Такого рода множественная устойчивость к лекарственным препаратам в настоящее время широко распространена среди многих патогенных микроорганизмов. Гены, придающие устойчивость к многим антибиотикам, соединены вместе в плазмидных факторах R (от англ. resistance - устойчивость), называемых также факторами устойчивости. Наиболее крупные из этих плазмид наряду с несколькими генами r содержат также фактор переноса устойчивости (resistance transfer factor - RTF) (рис. 31.14). Участок RTF позволяет плазмиде переноситься в другие бактерии с помощью конъюгации. В действительности гены участка RTF весьма сходны с аналогичными генами факторов F. Гены r кодируют ферменты, инактивирующие определенные лекарственные вещества. Факторы R, имеющие участок RTF, могут переноситься между различными видами бактерий при совместном культивировании. Следовательно, множественная устойчивость к антибиотикам может быть трансмиссивной.
Рис. 31.14. Схематическое изображение фактора R. Гены RTF (ответственные за конъюгацию и репликацию) показаны зеленым цветом, а гены г (ответственные за устойчивость к различным лекарственным веществам) — красным. IS-элементы показаны желтым цветом
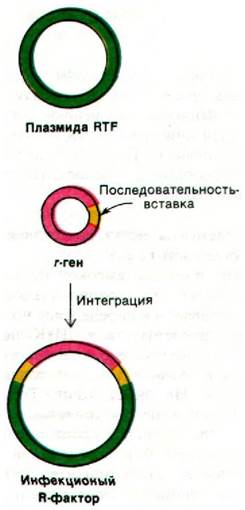
Маленькие плазмиды факторы R лишены области RTF и обычно придают клетке устойчивость только к одному антибиотику. Например, плазмида R pSC101 длиной 8,2 kb несет ген устойчивости к тетрациклину, но она не может быть перенесена путем конъюгации. Однако этот ген r может присоединиться к другой плазмиде, несущей иной ген устойчивости к какому-либо веществу (рис. 31.15). Если эти гены r интегрируют с плазмидой, содержащей область RTF, возникает трансмиссивная R-плазмида. Следовательно, плазмиды, являющиеся сложными факторами R, образуются из весьма подвижных элементов, обусловливающих, устойчивость к отдельным химическим соединениям. Такие генетические элементы, способные к переносу, теперь называют транспозонами.
Рис. 31.15. Инфекционный фактор R образуется, когда ген г присоединяется к плазмиде RTF
31.7. IS-элементы могут присоединяться к неродственным генам
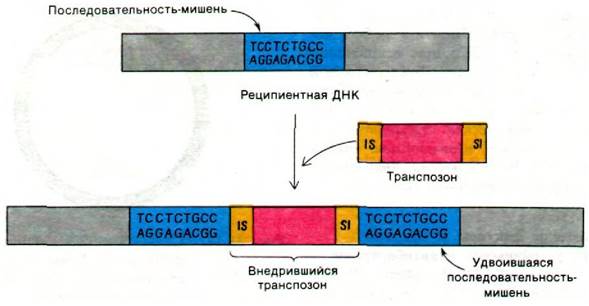
Что лежит в основе высокой подвижности транспозонов? Электронно-микроскопические исследования и определение последовательности нуклеотидов в ДНК показали, что последовательность, расположенная на одном конце транспозона, повторяется на другом конце. Например, концы Tn3-транспозона, кодирующего устойчивость к ампициллину, представляют собой обращенные повторы длиной 38 пар оснований. Последовательности нуклеотидов в ДНК рецепиенте, граничащие с обеих сторон с транспозоном, являются прямым повтором последовательности 5-9 пар оснований, присутствовавшей до вставки транспозона (рис. 31.16). Между этими обрамляющими последовательностями реципиентной ДНК и концевыми последовательностями транспозона нет никакой гомологии. Кроме того, гены rec E. coli не участвуют в процессе интеграции транспозона, что в корне отличает его от общей генетической рекомбинации. Концы транспозона, возможно, служат IS- элементами, направляя действие нуклеаз и других белков, участвующих в интеграции. Самое главное заключается в том, что транспозон не обязательно гомологичен реципиентной ДНК, так как специфичность интеграции определяется в первую очередь ДНК-белковыми взаимодействиями, а не спариванием оснований.
Рис. 31.16. Перенос гена (показано красным цветом), граничащего с обеих сторон с IS-элементами (желтый цвет). При транспозиции реципиентный участок (синий цвет) удваивается. Один конец транспозона представляет собой обращенный повтор другого конца
Самые маленькие подвижные генетические элементы - это последовательности- вставки (IS-элементы), которые имеют длину около 1 kb. В отличие от транспозонов IS-элементы не несут никаких генов. Однако они оказывают существенное влияние на выражение соседних генов. IS-элементы обычно блокируют транскрипцию дистальных генов транскрипционной единицы. Кроме того, они могут выступать в качестве новых промоторов. К тому же IS-элементы способствуют таким хромосомным перестройкам, как делеции и инверсии. В хромосоме E. coli было обнаружено несколько копий четырех различных IS-элементов (IS1, IS2, IS3 и IS4). К тому же концевые последовательности некоторых транспозонов идентичны одному из этих IS-элементов. По всей вероятности, транспозон образуется в том случае, когда какой-либо ген оказывается окруженным парой IS-элементов.
Мы уже видели, что плазмиды и фаги могут обмениваться блоками генов с бактериальными хромосомами. Кроме того, плазмиды и фаги способны рекомбинировать друг с другом. Было показано, что генетический элемент, отвечающий за устойчивость к тетрациклину, перемещается из плазмиды фактора R в фаг, размножающийся в клетках Salmonella, а оттуда в хромосому Salmonella, затем в фаг ![]() и из фага
и из фага ![]() в trp- оперон E. coli и обратно в фаг
в trp- оперон E. coli и обратно в фаг ![]() . Это замечательное путешествие показывает, насколько подвижны гены прокариот. Было бы интересно выяснить, обладают ли гены эукариот столь же высокой подвижностью.
. Это замечательное путешествие показывает, насколько подвижны гены прокариот. Было бы интересно выяснить, обладают ли гены эукариот столь же высокой подвижностью.