БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 31. ПЕРЕСТРОЙКИ ГЕНОВ: РЕКОМБИНАЦИЯ, ТРАНСПОЗИЦИЯ И КЛОНИРОВАНИЕ
31.11. Из суммарной геномной ДНК, расщепленной рестриктирующими эндонуклеазами, можно выделить с помощью клонирования определенные эукариотические гены
Как уже говорилось в предыдущей главе, исследование эукариотических геномов представляет огромные трудности. Ген длиной 1 kb составляет 2,5 • 10-4 генома Е. coli и всего лишь 3,4 • 10-7 генома млекопитающих. Методы рекомбинантных ДНК позволяют в настоящее время вводить эукариотический ген в Е. coli, что сильно упрощает задачу. Эксперимент начинается с частичного расщепления ДНК эукариотического генома, чтобы получить случайные фрагменты со средней длиной примерно 20 kb (рис. 31.25). К концам этих фрагментов присоединяют синтетические линкеры, образуют липкие концы и затем присоединяют к какому-нибудь вектору, например, к ДНК фага X. При упаковке ДНК в вирионы invitro происходит отбор рекомбинантных молекул ДНК, содержащих большие вставки. Затем этими рекомбинантными фагами заражают клетки Е. coli. Получается лизат, содержащий фрагменты эукариотической ДНК, заключенной в фаги и размноженной примерно в миллион раз. Этот лизат представляет собой библиотеку клонированной эукариотической ДНК.
Рис. 31.25. Стратегия клонирования, определенного эукариотического гена, начиная с расщепления суммарной геномной ДНК

Затем в такой библиотеке можно провести отбор для идентификации фаговых клонов, содержащих нужный эукариотический ген. Вычисление показывает, что лишь один из 180000 клонов будет содержать уникальный эукариотический ген. Поэтому необходима очень быстрая и эффективная процедура отбора. Она основана на гибридизации. Присутствие определенной последовательности ДНК в одной бляшке фага X можно выявить с помощью радиоактивной молекулы комплементарной ДНК или РНК в качестве гибридизационной пробы. Связывание этой пробы можно обнаружить методом радиоавтографии. Таким образом, можно проверить 1 млн. клонов за один день. Итак, клон, соответствующий определенному эукариотическому гену, можно легко идентифицировать и выделить при условии, что имеется транскрибированная с него РНК в более или менее чистом виде.
31.12. Эукариотические гены могут транскрибироваться в бактериальных клетках
Рекомбинантные молекулы ДНК, содержащие бактериальные гены, часто экспрессируются в клетках Е. coli. Например, клонирование триптофанового оперона Е. coli в составе плазмидного вектора ColE1 приводит к образованию больших количеств пяти биосинтетических ферментов, закодированных в этом опероне. Количество этих ферментов примерно в 20 раз выше, чем в обычной клетке Е. coli, так как рекомбинантная плазмида представлена многими копиями. ДНК дрожжей, простого эукариотического организма, также может экспрессироваться в бактериях. В одном исследовании в качестве клетки-хозяина использовали мутант Е. coli, нуждающийся в гистидине, так как он не содержал имидазолглицерол- фосфат-дегидрогеназы. При заражении этого мутанта фагом X, содержащим фрагмент дрожжевой ДНК, появилось несколько бактериальных клонов, которые уже не нуждались в экзогенном гистидине. Вставленный кусок дрожжевой ДНК нес недостающий ген, который экспрессировался с помощью механизмов транскрипции и трансляции бактериальной клетки.
Могут ли гены млекопитающих экспрессироваться в бактериях? Для ответа на этот вопрос в клетки Е. coli ввели ген инсулина крысы (рис. 31.26). Отправной точкой этого исследования послужила инсулинома - опухоль поджелудочной железы, выделяющая большое количество инсулина. Эта опухоль богата мРНК препроинсулина, предшественника активного гормона (разд. 35.9). С помощью обратной транскрипции этой мРНК была получена двухцепочечная комплементарная ДНК (кДНК), а затем ее включили в плазмидный вектор. Почему именно кДНК, а не геномную ДНК ввели в Е. соli? Как мы уже говорили, многие эукариотические гены содержат вставочные последовательности, которые вырезаются из первичных транскриптов (разд. 29.16). Поскольку бактерии, по всей вероятности, не способны удалять эти последовательности, представляется желательным трансформировать их участком ДНК, комплементарным зрелой мРНК. Действительно, оказалось, что несколько бактериальных клонов, трансформированных инсулиновой кДНК, синтезируют небольшое количество предшественника инсулина (около 100 копий на клетку). Недавно был получен еще один важный результат: последовательность ДНК, кодирующая овальбумин куриного яйца, экспрессируется в клетках Е. coli. Около 1,5% белка, синтезированного в таких трансформированных бактериях, представляет собой полные молекулы овальбумина (масса 43 кДа). Очевидно, гены эукариотических белков могут экспрессироваться в бактериях.
кДНК - комплементарная ДНК, синтезированная обратной транскриптазой на РНК- матрице.
Рис. 31.26. Синтез предшественника инсулина - проинсулина в трансформированных клетках Е. coli

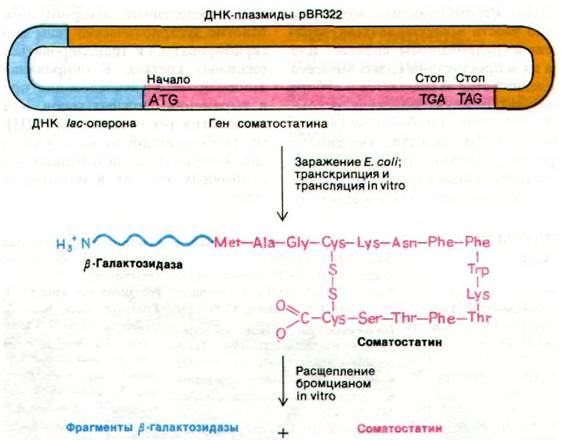
31.13. Химически синтезированный ген пептидного гормона соматостатина экспрессируется в клетках Е. coli
Последние достижения в химическом синтезе заданных последовательностей ДНК расширили масштаб и возможности метода рекомбинантных ДНК. Можно синтезировать de novo гены с практически любой нуклеотидной последовательностью и вставить их в вектор для введения в Е. coli. Прекрасным примером такого подхода служит синтез гена соматостатина - 14-членного пептида (рис. 31.27), который обнаруживается в экстрактах гипоталамуса. Соматостатин подавляет секрецию гормона роста, инсулина и глюкагона. Молекулу ДНК, кодирующую этот пептид, синтезировали путем соединения восьми олигонуклеотидных блоков. Этот ген соединили с геном ß-галактозидазы, локализованном в плазмидном векторе. При трансформации E. coli начался синтез гибридного белка, в котором соматостатин был присоединен к β-галактозидазе. Пептидную связь между этими двумя компонентами расщепляли in vitro с помощью бромциана (разд. 2.7). Для этого встроенный ген содержал кодон метионина перед первым кодоном соматостатина. С-конец соматостатина был свободен, так как после кодона последнего остатка пептида были расположены два стоп-кодона (рис. 31.27). На долю химерного белка приходилась значительная часть клеточного белка (около 3%). Более того, полученный таким способом соматостатин обладал биологической активностью. Следовательно, клонированием химически синтезированного гена можно получать функционально активный полипептид.
Рис. 31.27. Синтез пептидного гормона соматостатина клетками Е. соli, трансформированными химически синтезированным геном
31.14. Перспективы клонирования генов
Метод рекомбинантных ДНК открыл новые горизонты в молекулярной биологии. Увеличение числа генов путем клонирования в бактериях дает неограниченные количества ДНК для электронно-микроскопического анализа и определения последовательности нуклеотидов. Исследуются специфические участки ДНК, отвечающие за подвижность генов, репликацию ДНК и транскрипцию. Возникают совершенно новые направления; примером может служить открытие вставочных последовательностей во многих эукариотических генах. Появилась возможность быстро картировать сложные хромосомы и разделять их на отдельные элементы, которые поддаются различным манипуляциям. Темпы исследований постоянно нарастают. Кроме того, клонирование генов становится важным способом получения определенных белков в больших количествах. Например, с помощью рекомбинантных молекул можно увеличить в 500 раз количество ДНК-лигазы, образующейся в клетках Е. coli. Наиболее многообещающим представляется синтез пептидов и белков эукариот трансформированными бактериями. В недалеком будущем такие гормоны, как инсулин, и такие противовирусные агенты, как интерферон, будут продуцироваться бактериями1. В фармакологии начинается новая эра, которая, по-видимому, окажет глубокое воздействие на медицину. Исследуются также возможности клонирования генов для увеличения сельскохозяйственного производства. Эукариотические гены вводят в настоящее время не только в бактерии, но и в эукариотические клетки. Например, вирус SV-40 был использован в качестве вектора для переноса гена глобина кролика в клетки почек обезьяны. Такие зараженные клетки синтезировали значительные количества β-глобина кролика. Этот экспериментальный подход является многообещающим методом расшифровки регуляторных механизмов экспрессии эукариотических генов.
1 В Англии и США уже продается инсулин, синтезированный клетками бактерий. Прим. перев.