БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 32. ОБОЛОЧКИ БАКТЕРИАЛЬНЫХ КЛЕТОК
32.5. Синтез дисахарид-пептидного звена, прикрепленного к липидному переносчику
Следующий этап - присоединение NAG к остатку NAM в углеводпептидном звене, прикрепленном к липидному переносчику. Донор активированного углевода UDP- NAG вступает в реакцию с С-4 остатка NAM и между этими сахарами образуется β-1,4- гликозидная связь (рис. 32.9). Далее в ходе АТР-зависимой реакции NH4+ амидирует свободную α-карбоксильную группу D-глутаминовой кислоты в пептиде. Вслед за этим на ε-аминогруппе остатка лизина в пептиде выстраивается пентаглициновый мостик.
Рис. 32.9. Синтез дисахарид-пептидного звена на липидном переносчике
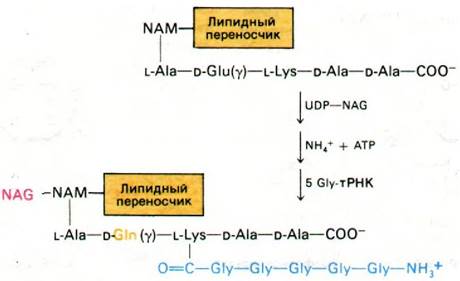
Этот пентапептид синтезируется путем последовательного присоединения глициновых остатков, доставляемых глицил-тРНК. Это единственный случай, когда тРНК служит донором аминокислот в ходе внерибосом- ного синтеза пептида. На этом завершается образование основной структурной единицы клеточной стенки.
32.6. Перенос дисахарид-пептидного звена на растущую полисахаридную цепь
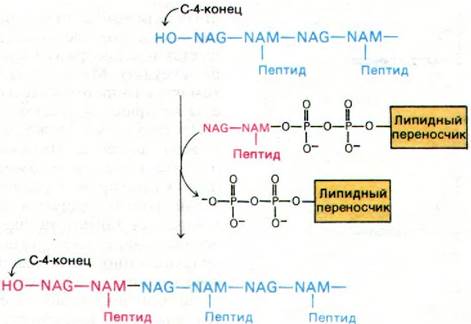
Дисахарид-пептидное звено переносится липидным переносчиком на нередуцирующий конец растущей полисахаридной цепи. Реакция протекает таким образом. Атом углерода С-1 остатка NAM находится в активированном состоянии, поскольку он связан с липидным переносчиком пирофосфатной связью. Поэтому он вступает в реакцию с С-4 концевого остатка NAG растущей полисахаридной цепи, образуя в итоге β-1,4- гликозидную связь (рис. 32.10).
Рис. 32.10. Перенос дисахарид-пептидно- го звена на растущую полисахаридную цепь
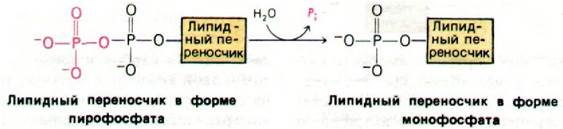
Липидный переносчик высвобождается в форме пирофосфата и далее гидролизуется до монофосфата специфической фосфатазой. Этот этап дефосфорилирования представляет собой, по существу, регенерирование переносчика, способного акцептировать углеводпептидное звено от UDP. Известен пептидный антибиотик бацитрацин, ингибирующий биосинтез клеточных стенок путем торможения этого этапа:
32.7. Поперечные мостики между полисахаридными цепями образуются в реакции транспептидирования
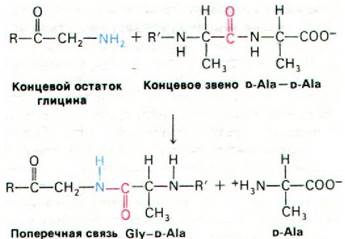
В результате реакции транспептидирования между полисахаридными цепями образуются поперечные сшивки, что приводит к формированию одной огромной молекулы, напоминающей по виду мешок. В ходе транспептидирования концевая аминогруппа одного пентаглицинового мостика атакует пептидную связь между остатками D-Ala-D-Ala другой пептидной единицы (рис. 32.11). При этом образуется пептидная связь между глицином и одним из остатков D-аланина, а второй остаток D-аланина высвобождается. Фермент, катализирующий эту реакцию, - гликопептид-транспептидаза. Обратите внимание, что синтез этой поперечной связи не требует расхода АТР: реакция идет за счет свободной энергии, уже содержащейся в связи D-Аlа-D-Аlа. Формирование пептидной связи таким необычным способом определяется, очевидно, тем, что реакция протекает вне клетки, т.е. в остсутствие АТР. Вспомните, что путем трансаминирования без использования АТР образуются пептидные связи при поперечной сшивке нитей фибрина (разд. 8.21).
Рис. 32.11. Аминогруппа пентаглицинового мостика атакует пептидную связь между двумя остатками D-Ala; в результате формируется поперечная связь (сшивка)
32.8. У грамположительных бактерий пептидогликан покрыт тейхоевой кислотой
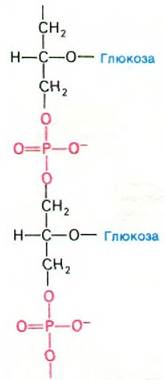
Поверхность грамположительных бактерий состоит из тейхоевой кислоты, которая представляет собой полимер остатков глицерола (или другого углевода, например, рибитола), соединенных фосфодиэфирными мостиками (рис. 32.12). Свободные гидроксильные группы этерифицируются с аланином или сахарами, в частности с глюкозой. Тейхоевая кислота присоединяется к скелету пептидогликана - последовательности остатков NAG-NAM - фосфодиэфирной связью. Удлинение цепей тейхоевой кислоты происходит путем переноса глицерол- фосфата от CDP-глицерола на свободную концевую ОН-группу цепи. Остатки глюкозы присоединяются к гидроксильным группам скелета тейхоевой кислоты в ходе реакции с UDP-глюкозой.
Рис. 32.12. Строение тейхоевой кислоты