Структура и функционирование белков. Применение методов биоинформатики - Джон Ригден 2014
Предсказание функции белков на основе их теоретических моделей
Практическое применение
Предсказания функции на основе моделей ab initio
До недавнего времени, пока не были разработаны более мощные, но ресурсоёмкие алгоритмы (Bradley et al. 2005), разумной целью моделирования без использования шаблона (ab initio или de novo) было лишь получение правильной укладки, нежели более точные предсказания (см. Главу 1). Это ограничивало диапазон применимых методов предсказания функции белков, и означает, что большинство предсказаний, описанных в литературе, основаны, главным образом, на предсказаниях укладки белка и её корреляции с его функцией (обсуждалось в Главе 6).
В одном из первых масштабных применений сервера ROSETTA, Бонно с коллегами (Bonneau et al., 2002) создал модели для 510 белковых семейств по классификации Pfam со средней длинной менее 150 остатков. Это были неизвестные к тому времени структуры белков, но для некоторых из них функция была известна или предполагалась. В нескольких случаях умозрительные предсказания могли быть подкреплены результатами моделирования. Например, предполагали, что PF01938, домен белка TRAM, может связываться с нуклеиновыми кислотами. Это предположение строилось, в основном, на сходстве его ab initio модели со структурами в надсемействах базы данных SCOP, содержащих различные белки, связывающие нуклеиновые кислоты. Теперь мы можем судить о точности модели по двум неопубликованным результатам проекта по структурной геномике (lyez и lyvc). Примером предсказания функции для совершенно не охарактеризованного белка служит домен с неизвестной функцией 37 (PF01809). Его модель подошла в качестве структуры NK-лизина - гемолитического белка, экспрессируемого в естественных киллерах. Хотя структура белка PF01809 остается неизвестной, база данных Pfam на момент написания этих строк сообщает неопубликованные свидетельства, что такой белок из Aeromonas hydrophila действительно демонстрирует гемолитическую активность.
Примечательно, что одна ab initio модель не обязана в точности соответствовать известному типу укладки, чтобы дать ключ к пониманию функции; напротив, иногда на решение наталкивает весь широкий класс структур, которому принадлежит конкретная модель. Примером этого является модель, построенная для муцин-связывающего домена (Bumbaca et al. 2007). Предпочтительная модель содержала укладку типа “бета- сэндвич” такого типа, который сильно коррелирует со связыванием углеводов, - на момент публикации половина семейств углевод-связывающих доменов с известной структурой имела такую укладку. Это скорее могло бы согласовываться со связыванием домена с углеводородной частью его мишени, высокогликозилированного муцина, нежели чем с его белковой частью. Более того, эта ab initio модель имела три экспонированных ароматических остатка такого типа, который считается характерным для связывания углеводородов (Quiocho and Vyas 1999).
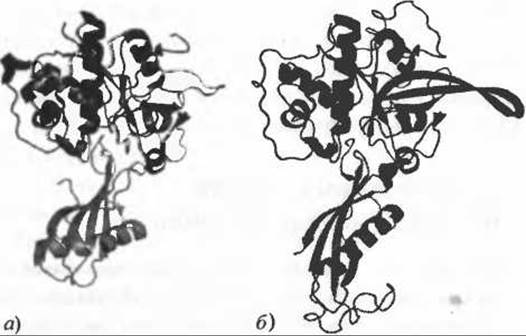
Рис. 12.2. Предсказание белок-белковых взаимодействий на основе моделирования структуры белков. Подход, основанный на сравнительном моделировании структуры белковых комплексов (Davis et al. 2008). позволил, используя структуру катепсина А в комплексе со стефином A (PDB код 1nb3; а) предположить возможное взаимодействие между фальципаином-2 и цистатином, что и было подтверждено кристаллографически (PDB код 1yvb; б). На обеих панелях ферменты показаны сверху, ингибиторы снизу
Недавний пример показал, как функция, предсказанная по укладке ab initio модели может быть подтверждена другими методами (Rigden and Galperin 2008). Известно, что белок SpoVS необходим для спорообразования у спорообразующих бактерий, но на самом деле распространен гораздо шире. Фенотипическое описание организмов, имеющих мутированный SpoVS, мало что говорит о его молекулярной роли. Однако лучшие модели, полученные с помощью серверов ROSETTA и I-TASSER, хорошо соответствуют укладке белка Alba, содержащегося в хроматине архей (Рис. 12.3а). Такая укладка тесно связана со связыванием нуклеиновых кислот в различных случаях и, более того, картирование электростатического потенциала на поверхности моделей выявило отчетливую положительно заряженную область характерную для белков, связывающихся с нуклеиновыми кислотами (Рис. 12.3b; см. Главу 7). Суммируя результаты этих методов можно предположить, что белок SpoVS является новым фактором транскрипции, который вовлечен в контроль запутанной схемы экспрессии генов, происходящей при споруляции (Rigden and Galperin 2008).
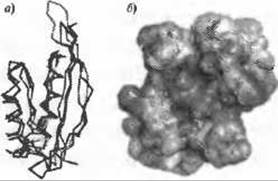
Рис. 12.3. (Цветную версию рисунка см. на вклейке.) Анализ ab initio моделей белка SpoVS позволил предположить наличие у него функции связывания нуклеиновых кислот (Rigden and Galperin 2008). а) Модели, полученные как с помощью ROSETTA (показано серым) так и с помощью I-TASSER (показано черным) сильно схожи со структурой белка Alba, содержащемся в хроматине архей (PDB код 1nfj; окрашен по спектру от синего N-конца до красного С-конца). б) Электростатический потенциал возможного димера SpoVS, построенного по модели ROSETTA (синим показаны области положительного потенциала, красным - отрицательного)
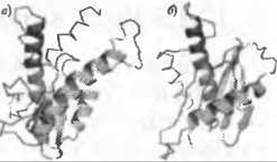
Рис. 12.4. (Цветную версию рисунка см. на вклейке.) Подтвержденное предсказание структуры, выполненное Мальмстрем с коллегами (2007). Модель белка TRS20/YBR254C (а) соответствовала надсемейству SNARE по классификации SCOP; это соответствие позже было подтверждено получением экспериментальной структуры (PDB код 1h3q) схожего белка (б). Цветом выделены структурно схожие элементы, остальное показано серым
Недавнее масштабное применение ab initio моделирования в рамках подхода, включавшего также использование PSI-BLAST и методов предсказания структуры, основанных на протягивании, было направлено на анализ генома дрожжей (Malmstrom et al. 2007). Авторы применили новую стратегию по использованию известной информации о функции, чтобы облегчить отбор правильных совпадений между потенциальными структурами ab initio моделей и надсемейств по SCOP. С этой целью вдобавок к сравнению структур было оценено совпадение по классификации ГО между белком-мишенью и белками рассматриваемых надсемейств. Эти дополняющие друг друга источники информации были объединены с применением байесовской статистики. На рисунке (Рис. 12.4) показан пример предсказания принадлежности белка TRS20/YBR254C к надсемейству SNARE по классификации SCOP, которое позже было подтверждено экспериментальной расшифровкой структуры. Совпадение между моделью и кристаллографической структурой частичное и умеренное (Рис. 12.4), что иллюстрирует ценность информации, относящейся к белку-мишени и надсемействам потенциально совпадающих структур, которая содержится в БД ГО. В рассматриваемом случае мишень TRS20/YBR254C являлась одной из субъединиц комплекса переноса белковых частиц (англ. transport protein particle (TRAPP)), участвующем в состыковке и слиянии везикул. Ee совпадение со структурой из надсемейства SNARE по классификации SCOP было, таким образом, надежно подтверждено, поскольку везикулярный транспорт является одной из главных функций белков этого надсемейства.