БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
УТВОРЕННЯ ЖИВИХ СИСТЕМ ТА ЇХ СТРУКТУРНА ОРГАНІЗАЦІЯ
ХАРАКТЕРИСТИКА ОСНОВНИХ СТРУКТУРНИХ КОМПОНЕНТІВ КЛІТИНИ
Для різних видів клітин рослинного і тваринного світу важливими структурними компонентами є системи мембран.
Мембранна система клітин охоплює зовнішню плазматичну мембрану, а також складний комплекс внутрішніх мембран (ендомембран) і мембран клітинних органел — мітохондрій, пластид і у евкаріотів — клітинного ядра. Кожна мембрана є досить складним утворенням з характерними структурою, складом і певними функціями.
Товщина мембран становить всього 6—12 нм, але вони мають високу міцність, стійкість, гнучкість і лабільність. За цими показниками мембрана перевищує багато природних, штучних і синтетичних матеріалів, які застосовуються в техніці.
Загальна площа мембран в органах і тканинах організму досить велика. У середньому на 1 г білка ендоплазматичної сітки печінки припадає 50 м2 мембран. Така велика площа мембран, незважаючи на мікроскопічні розміри клітини, дає можливість ефективно забезпечувати різноманітні обмінні процеси.
Мембрани різних клітин і органів дещо відрізняються між собою як за хімічним складом, так і за відносним вмістом компонентів, що відіграє важливу роль у забезпеченні широкого діапазону різноманітних видів фізіологічної активності. Однак, незалежно від своєї морфологічної будови і хімічного складу, мембрани є досить ефективним засобом локалізації процесів всередині клітини.
Кожна мембрана відокремлює цитоплазматичний простір від неплазматичного, бере участь в утворенні компартментів клітини — ділянок з різноманітною метаболічною активністю, забезпечуючи одночасний перебіг в одній клітині різних біохімічних реакцій. Розділяючи клітину на велику кількість ізольованих ділянок, мембрани забезпечують збереження в кожній з них специфічних умов і фізико-хімічних показників, таких, як pH розчину, температура, концентрація розчинених речовин, електричний потенціал.
Мембрани клітин—досить динамічні системи, у клітині вони перебувають в постійному русі. Мембрани дуже швидко руйнуються і легко відновлюються над пошкодженими ділянками, зливаються одна з одною, розтягуються і стискуються під час руху та внаслідок зміни форми клітин.
У життєдіяльності клітин мембрани відіграють надзвичайно важливу роль. Вони регулюють велику кількість метаболічних процесів як всередині клітини, так і на її поверхні. Мембрани беруть участь у формуванні структури клітини і клітинних органел. Вони оточують ядро, формують тіло мітохондрій, структуру хлоропластів, утворюють ендоплазматичний ретикулум і апарат Гольджі. Завдяки своїм фізико-хімічним, біологічним і структурним особливостям мембрани виконують цілий ряд важливих функцій, таких, як регуляція транспорту молекул та іонів, забезпечення специфічної рецепції, «пізнавання» клітин, визначають антигенну специфічність.
Беруть участь мембрани і в енергетичних процесах клітини, у передачі нервових імпульсів, забезпеченні фотосинтетичних реакцій. Завдяки субмікроскопічній структурі між зовнішньою і внутрішньою сторонами мембран утворюється, формується і підтримується на певному рівні різниця електричних потенціалів, що відіграє важливу роль у забезпеченні ряду метаболічних реакцій. Значно впливають мембрани на різноманітні метаболічні процеси всередині клітини, оскільки вони координують деякі ферментативні реакції. У мембранах міститься ціла система різноманітних ферментів, кожний з яких перебуває в чіткій взаємодії з іншими. Крім того, мембрани значною мірою змінюють активність різних ферментів. Деякі ферменти активні лише тоді, коли перебувають у тісному контакті з мембранами, а інші виявляють максимум каталітичної активності в тому разі, коли звільняються і переходять у цитоплазму. В організмі є велика кількість мембран, які, крім основних функцій, виконують ще й спеціальні — всмоктування і перетравлювання їжі, скорочення і розслаблення м’язів, трансформацію різних подразників в електричні імпульси та ін.
Основними структурними компонентами більшості мембран є білки, ліпіди і вуглеводи. Кожний з цих структурних компонентів має певні характерні особливості будови і властивості, що зумовлюють участь їх у побудові клітинних структур.
Першу модель біологічної мембрани розроблено в 30-х роках американськими дослідниками. За цією моделлю білкові молекули розміщуються на зовнішньому і внутрішньому боці бімолекулярного ліпідного шару. Таким чином створюється структура білково-ліпідного бутерброда — зверху і знизу розміщені білкові молекули, а посередині — ліпіди. Така бутербродна модель мембрани тривалий час вважалась універсальною для всіх живих систем. Та з появою нових методів дослідження було встановлено ряд нових фактів, які не вдавалось пояснити з погляду мембранної структура Тому поступово бутербродна модель мембрани була відкинута і запропонована рідинно-мозаїчна модель, яка зараз є загальноприйнятою.
За сучасними уявленнями клітинна мембрана є гетерогенною структурою, що складається з двомірно орієнтованих розчинів глобулярних білків та їх агрегатів (кластерів) у бімолекулярному шарі складних ліпідів. Часто мембрани зображують у вигляді ліпідного «моря», по якому «плавають» білкові «айсберги» (мал. 3).
Структура і функції мембранних ліпідів. Основними компонентами біологічних мембран є молекули складних ліпідів, в яких досить виражена структур на асиметрія. Ліпіди складаються з двох частин: полярної головки, що несе електричні заряди, і неполярного хвоста.
Полярна частина становить не більше як V* молекули і містить у своєму складі фосфати, цукри, холін, коламін, що зумовлюють наявність заряду. У переважній більшості полярні головки ліпідів несуть негативний заряд або є електронейтральними.
Неполярна частина молекули містить в своєму складі незаряджені залишки жирних кислот та інших сполук. Зв’язуючою ланкою між полярною і неполярною частинами молекул здебільшого є залишок гліцерину (гліцероліпіди) або аміноспирту сфінгозину (сфінголіпіди). Неполярні хвости ліпідів відрізняються один від одного ступенем насиченості, розгалуженістю і кількістю вуглецевих атомів. Наявність подвійних зв’язків у вуглеводних ланцюгах неполярних хвостів ліпідних молекул має важливе значення для нормального функціонування біологічних мембран живих систем. Було виявлено, що від ступеня насиченості мембранних ліпідів залежить швидкість проникнення різних речовин через біологічні мембрани.
Крім фосфоліпідів, до складу багатьох мембран, особливо тваринних клітин, входять також гліколіпіди і холестерин. Особливо велика кількість холестерину міститься в зовнішніх мембранах, зокрема в плазматичних мембранах клітин печінки, де вміст його становить приблизно 30 %.
Вміст ліпідів у складі мембран неоднаковий і залежить від їх виду і функцій. Мембрани з високим вмістом ліпідів формуються там, де потрібно створити чітку межу розподілу з навколишнім середовищем — на поверхні клітин, ендоплазматичного ретикулуму, вакуолей. Формування ліпідних шарів у мембранах є біологічно доцільним при несприятливих умовах у клітині, у випадку формування ізолюючих (діелектричних) прошарків на шляху руху електронів.
Структура і функції мембранних білків. Молекули мембранних білків у переважній більшості являють собою об’ємні структури — глобули і подібно до ліпідів характеризуються структурною асиметрією. Завдяки наявності в складі поліпептидних ланцюгів амінокислот з полярними і неполярними радикалами в водному середовищі молекула білка орієнтується таким чином, що неполярні ділянки її, уникаючи контакту з водою, розміщуються всередині глобули, а полярні — на її поверхні. Частини молекул, розміщені на поверхні глобул, виступають з мембран і мають фіксовані заряди. На зовнішній поверхні сумарний заряд виявляється негативним.
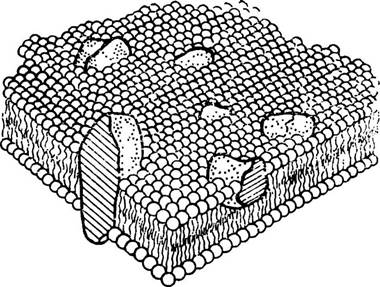
Мал. 3. Мозаїчна модель клітинної мембрани.
Усі мембранні білки беруть участь у дифузійному русі, тому їх розподіл на поверхні мембрани в певний момент є випадковим. Вважають, що білки плавають у ліпідному шарі подібно до айсбергів. Однак швидкість такого переміщення різна для різних білків і досить незначна. Вона залежить не тільки від в’язкості мембран, а й від глибини занурення білків у ліпідний біошар.
Вміст білків у різних мембранах неоднаковий і залежить від функцій, які виконуються ними. Високий вміст білків у мембранах, в яких відбуваються численні ферментативні реакції. Так, внутрішні мембрани мітохондрій містять більш як 75 % білків. Високий вміст білків (50 %) також у цитоплазматичній мембрані. У мембранах клітин мієлінових оболонок, які виконують в основному роль ізоляторів, вміст білків становить лише 20 % від загальної маси мембран.
Залежно від вмісту білків у складі мембран відбувається і їх розподіл. Молекули білків розподіляються в ліпідній структурі у вигляді мозаїки, якщо їх небагато. А коли вони становлять значний вміст, то набирають форми дірчастої сітки, петлі якої заповнені ліпідами.
Плазматична мембрана. Для нормального функціонування рослинних і тваринних організмів важливе значення має наявність своєрідної клітинної межі — плазматичної мембрани, яка забезпечує регуляцію обміну речовин між клітиною і зовнішнім середовищем. Товщина плазматичної мембрани в переважній більшості становить 6— 8 нм, тому її можна спостерігати лише в електронний мікроскоп. Хімічний склад плазматичної мембрани рослинних і тваринних організмів практично однаковий. До її складу входять білки, ліпіди, полісахариди, ферменти та різні іони, тобто всі групи сполук, з яких складається клітина.
Плазматична мембрана є своєрідним регулятором метаболічних процесів у клітині і має важливе значення для організму в цілому. На ній відбуваються найрізноманітніші біохімічні реакції, від яких залежить функціонування організму як складної високоорганізованої живої системи. Мембранні компоненти беруть безпосередню участь у транспорті різних речовин, молекул та іонів у клітині, де відбувається їх утилізація. Ці процеси часто супроводжуються синтезом макроергічних сполук та виділенням енергії, що необхідна для забезпечення метаболічних процесів.
З участю плазматичної мембрани відбуваються процеси специфічної рецепції, міжклітинної взаємодії та передачі інформації між окремими клітинами, органами і системами організму.
У плазматичній мембрані міститься набір специфічних ферментів і рецепторів, природа яких визначається особливостями будови і функціями клітин.
Оскільки клітини для забезпечення процесів життєдіяльності асимілюють речовини із зовнішнього середовища, досить важливою є також бар’єрна функція плазматичної мембрани — здатність вибірково пропускати одні речовини і обмежувати пропускання інших.
Плазматична мембрана клітин — неоднорідне утворення. Вона складається з різних ділянок, які відрізняються одна від одної як за молекулярною структурою, так і за хімічним складом. На деяких ділянках плазматична мембрана має гладку поверхню, на інших може бути покрита мікроворсинками, мати різноманітні вигини залежно від функцій, що їх виконує клітина. Ці утворення можуть бути тимчасовими і виникають за певних умов (під час вбирання чи виділення певних речовин) або існують постійно.
Плазматичні мембрани спеціалізованих клітин іноді мають своєрідну структуру. Так, у клітинах кишкового епітелію плазматична мембрана утворює подібні вгинання — мікроворсинки, завдяки яким значно збільшується площа клітинної поверхні і підвищується ефективність обміну речовин між внутрішнім середовищем (цитоплазмою) і позаклітинним простором. Глибокі вгинання плазматичної мембрани ниркових канальців (базальний лабіринт) виконують важливу секреторну функцію.
У бактеріальних клітинах внаслідок зливання увігнутих ділянок мембрани утворюються кармани, в яких містяться пластинчасті, трубчасті чи везикулярні тільця — мезосоми. Внутрішній простір мезосом частково стикається з позаклітинним простором. Функції мезосом повністю не з’ясовано, вважають, що вони є місцем прикріплення ДНК ядерної зони.
Значно впливають на поверхневі властивості мембрани та її функції і життєдіяльність зв’язані з мембраною екстрацелюлярні компоненти. У багатьох випадках ці утворення можна спостерігати лише під електронним мікроскопом. Це, зокрема, екстрацелюлярне утворення, що міститься на поверхні тваринних клітин і називається глікокаликсом.
Глікокаликс містить складні вуглевод-білкові комплекси — глікопротеїди. Розміщуються глікопротеїди на поверхні мембрани таким чином, що С-кінець поліпептидного ланцюга виступає з внутрішнього боку мембрани, а N-кінець з численними розгалуженими боковими ланцюгами полісахаридів міститься на поверхні мембрани.
З глікопротеїдних компонентів глікокаликсу досить вивченим є глікофорин мембран еритроцитів, який забезпечує антигенну специфічність та зв’язування різних вірусів і лек- тинів. До складу глікофорину входить близько 60 % вуглеводів. Функції глікокаликсу досить різноманітні. Вважають, що він відіграє важливу роль у процесах рецепції, пізнавання та міжклітинної взаємодії. Яйцеклітини більшості тварин містять на поверхні особливий тип глікокаликсу — віте- лінову оболонку. У ссавців, крім того, є товста прозора зовнішня оболонка, відокремлена від плазматичної мембрани простором, що заповнений рідиною (перивітеліновий простір), яка містить вуглеводи, білки, глікопротеїди. Вітеліно- ва оболонка відіграє важливу роль у забезпеченні специфічних функцій даних клітин — на її поверхні містяться специфічні рецептори, що зв’язують сперматозоїди.
На поверхні плазматичної мембрани багатьох клітин часто містяться також інші екстрацелюлярні речовини, що є продуктами життєдіяльності цитоплазми (слизі, хітин, мінеральні солі), більшість з яких відіграє важливу роль у процесах життєдіяльності клітини. Слизові виділення багатьох водоростей забезпечують процеси гідратації, жорсткі хітинові покриви комах виконують захисні функції. За рахунок екстрацелюлярних компонентів забезпечується також еластичність, опірність та адгезійна здатність клітин, без чого існування багатоклітинних організмів було б неможливим. У бактерій зовнішній шар клітинної оболонки визначає також імунологічний тип, вірулентність.
Рослинні клітини, крім плазматичної мембрани, мають добре виражену відносно товсту клітинну стінку — целюлозну оболонку. Вона утримує протопласт і надає йому певної форми. Тому більшість рослинних і бактеріальних клітин, на відміну від тваринних, не можуть змінювати своєї форми.
Мембранний транспорт. Досить важливою є роль плазматичної мембрани в забезпеченні бар’єрної функдії, яка полягає в регулюванні надходження різних речовин у клітину та виходу їх у міжклітинний простір. У нормальних фізіологічних умовах між клітиною і навколишнім середовищем відбувається постійний обмін різними речовинами і іонами, тому мембрани не можуть бути повністю непроникними. Навіть у тому випадку, коли мембрана перебуває в стані спокою і канали її закриті, частина речовин може дифундувати крізь неї. Так, крізь мембрану легко проходять молекули води, невеликі молекули інших речовин і деякі іони.
Плазматична мембрана має осмотичні властивості. Осмос— це однобічна дифузія розчинника в розчин крізь напівпроникну мембрану. Осмос спостерігається тоді, коли напівпроникна мембрана розділяє два розчини різної концентрації, розчинник при цьому переміщуватиметься від розчину меншої концентрації до розчину більшої концентрації. Якщо помістити клітини в чисту воду, вона за градієнтом водного потенціалу (φ) проникатиме всередину клітини. В результаті осмотичного вбирання води збільшується об’єм клітин і відбувається їх руйнування (гемоліз). Так поводяться клітини в розчинах, осмотичний тиск яких менший від осмотичного тиску клітинного вмісту (гіпотонічні розчини). У розчинах, осмотичний тиск яких більший від осмотичного тиску внутріклітинного вмісту (гіпертонічні розчини), відбувається плазмоліз, зморщування клітин внаслідок виходу води з клітини.
Розчини, осмотичний тиск яких дорівнює осмотичному тиску клітинного вмісту, називаються ізотонічними, вони не впливають на зміну об’єму клітинного вмісту і застосовуються для введення в кров і міжклітинну рідину при різних порушеннях обміну.
Різні речовини можуть проникати крізь цитоплазматичну мембрану внаслідок пасивного перенесення, за градієнтом концентрації або внаслідок полегшеної дифузії, з участю мембранних переносників.
Транспорт крізь плазматичну мембрану порівняно великих частинок і крапель рідини відбувається з участю ендо- і екзоцитозу.
Механізм мембранного транспорту різних речовин вивчено ще недостатньо. Вважають, що пасивний транспорт забезпечується за рахунок кінетичної енергії речовин, які переносяться. При цьому найлегше відбувається транспорт ліпофільних речовин, які мають здатність розчинятися в мембранних ліпідах і переносяться в розчині за законами дифузії. Пасивний транспорт гідрофільних речовин відбувається крізь субмікроскопічні нори (тимчасові пустоти), які є нестабільними структурними утвореннями, що виникають в результаті тимчасової локальної перебудови молекулярної організації динамічної мембрани.
Транспорт заряджених молекул і іонів залежить не тільки від градієнта концентрації, а й від різниці потенціалів. Причому дифузія іонів відбувається в напрямі електрохімічного потенціалу.
Численні експерименти показали, що проникність мембран для різних молекул та іонів значною мірою залежить від специфічних канальних (тунельних) білків. Ці білки беруть безпосередню участь в утворенні каналів, через які проходять катіони, аніони чи незаряджені молекули.
Перехід через мембрану речовин, для яких вона є практично непроникною (амінокислот, цукрів, нуклеотидів), відбувається за механізмом полегшеної дифузії, з участю специфічних транспортних білків. Речовини, для яких характерна полегшена дифузія, як правило, добре розчинні у воді і нерозчинні в ліпідах. При утворенні комплексу речовина-переносник дещо змінюються властивості транспортованих речовин, і вони набувають здатності добре розчинятися в ліпідах мембран.
Переносники відрізняються один від одного як за механізмом дії, так і за субстратною специфічністю. За механізмом дії переносники поділяють на дві групи. До першої групи належать переносники, які діють за човниковим механізмом — транспортують через мембрану певні речовини, після чого комплекс речовина-переносник розпадається і цикл знову повторюється. Інколи переносник у зворотньому напрямі може транспортувати інші речовини.
До другої групи належать переносники, вбудовані в структуру мембран. Протомери цих олігомерних білків-переносників утворюють гідрофільний канал-пору з клапаном, через яку ніби по естафеті від молекули до молекули передають транспортовану речовину. Перенесення речовин через мембрану відбувається за рахунок зміни конформації ділянок, зв’язаних із субстратом, внаслідок чого в мембрані відкриваються канали, по яких відбувається транспорт.
Транспорт речовин з участю переносників часто залежить від присутності інших речовин, відбувається так званий зв’язаний транспорт. Коли один переносник транспортує два різних субстрати в одному напрямі, це називається паралельним транспортом — симпортом. Прикладом може бути перенесення протонів і лактози у кишкової палички. Якщо транспорт одних речовин пов’язаний з перенесенням
інших у протилежному напрямі, відбувається антипаралельний спряжений транспорт — антипорт. Прикладом може бути транспорт в протилежних напрямах іонів Na+ і К+ через плазматичні мембрани, перенесення АТФ і АДФ через внутрішню мембрану мітохондрій та ін.
Активний транспорт молекул та іонів через мембрану. Для більшості клітин рослинних і тваринних організмів характерним є різний іонний склад з обох боків плазматичної мембрани. Так, всередині клітини переважають іони К+ і значно нижча концентрація іонів Na+, а в позаклітинному просторі навпаки. Отже, К+ — інтрацелюлярні іони, a Na+ — екстрацелюлярні. І хоч плазматична мембрана обмежує пасивне проникнення іонів, дифузні потоки у разі відсутності належної протидії з часом зрівняли б концентрацію та іонний склад всередині клітини і в позаклітинному просторі. Тому для підтримування необхідної концентрації іонів, крім вільної і полегшеної дифузії, існує механізм, який забезпечує активний транспорт іонів проти електрохімічного градієнта з участю так званих Na+—К+ і Na+—Са2+-насосів. Оскільки перекачування іонів відбувається проти градієнтів концентрації, то даний процес є енергозалежним і пов’язаний з екзергонічними реакціями (реакціями, що є джерелом енергії).
Основна функція Na+—К+-насоса полягає в тому, що він забирає з клітини іони Na+ і обмінює їх на іони К+. При цьому в середньому на кожних три іони Na+ обмінюється два іони К+ Оскільки перехід іонів здійснюється проти градієнта концентрації, то даний процес може відбуватися лише з використанням енергії АТФ. На один цикл перенесення витрачається одна молекула АТФ.
Функціонування натрій-калієвого насоса має надзвичайно важливе значення для метаболічних процесів організму. «Відключення» даного насоса на 5—10 хв в ізольованих нейронах приводить до зміни іонного складу на 40—50 %. Тому, коли порушується діяльність біологічних насосів, клітини гинуть.
Робота насоса регулюється іонами Na+ і К+. З внутрішнього боку мембрани міститься центр зв’язування іонів Na+, а з зовнішнього — іонів К+. Одночасне зв’язування цих іонів приводить насос в активний стан. У здійсненні цього процесу бере участь фермент (Na+, К+) -залежна АТФ-аза. Процес перенесення іонів поділяється на кілька стадій. Спочатку в присутності АТФ та іонів Mg2+ з внутрішнього боку мембрани фермент захоплює іон Na+ і переносить його в позаклітинний простір. Молекула АТФ при цьому передає ферменту макроергічний зв’язок і перетворюється в АДФ. Після перенесення іона Na+ з участю ферменту та енергії АТФ макроергічний зв’язок внаслідок втрати енергії знову перетворюється в простий, при цьому АТФ-аза захоплює іон К+ і переносить його всередину клітини. За цих умов здійснюється синтез АТФ — передача макроергічного зв’язку з АТФ-ази на АДФ. Важливою особливістю Na+— К+-насоса є зворотність дії його і залежність коефіцієнта зв’язаності іонів Na+ і К+ від протилежно спрямованих градієнтів. Зворотність дії насоса підтверджується тим, що існує певний енергетичний бар’єр, при досягненні якого виведення іонів Na+ з клітини припиняється.
Іонний транспорт, що забезпечується біологічними насосами, відіграє важливу роль у використанні хімічної енергії та перетворенні її в електричну. Якщо швидкість виходу з клітини іонів Na+ більша від швидкості надходження іонів К+, між внутрішнім і зовнішнім боками мембрани виникає різниця потенціалів, яка забезпечує протікання ряду метаболічних процесів — передачу нервових імпульсів, скорочення м’язів тощо.
Здобуто дані, які свідчать, що в функціонуванні Na+— К+-насоса важливу роль відіграють специфічні канальні білки мембран, що їх утворюють натрієві і калієві канали, розміщені поряд. При зв’язуванні іонів Na+ змінюється конформація білкових молекул, порушується система водневих зв’язків і відбувається перехід α-спіралі в рихлішу α-спіраль, в якій на один виток припадає не 3,6 амінокислотного залишку, а 4,4. Внаслідок цього утворюється внутрішній канал, який пропускає іон Na+, але не достатній для проходження іона К+. Після проходження іона Na+ знову відбувається зміна конформації і перетворення π-спіралі в α-спіраль. При цьому натрієвий канал закривається, а стінки сусіднього — калієвого — розходяться і пропускають іони К+.
Нині активно вивчається інший іонний насос — Na+— Са2+, який забезпечує взаємний обмін іонів Са2+ на іони Na+. При цьому відбувається нагнітання іонів Са2+ в напрямі, протилежному градієнту їх концентрації. У кожному циклі відбувається транспорт всередину клітини іонів Са2+ і вихід іонів Na+ або навпаки — вихід з клітини іонів Са2+ і перехід в клітину іонів Na+ з позаклітинного простору. Функціонування насоса забезпечується білком-переносником, який утворює мембранний канал і з участю АТФ-ази здійснює транспорт іонів.
Роль кальцієвого насоса полягає в забезпеченні певної концентрації іонів Са2+ в клітині, оскільки вони мають важливе значення для запускання ряду внутріклітинних процесів. Так, збільшення концентрації іонів Са2+ всередині клітини є сигналом для скорочення м’язових волокон. Іони Са2+ ініціюють процеси клітинного поділу, руху, секреції ряду біологічно активних речовин тощо.
Плазматична- мембрана не тільки відіграє важливу роль у регулюванні проникнення в клітину молекул та іонів, а й забезпечує вбирання твердих і рідких речовин на основі піноцитозу і фагоцитозу. З фізіологічного погляду ці два процеси рівнозначні і забезпечують проникнення в клітини значних за розміром речовин, не здатних самостійно проходити через плазматичну мембрану.
Термін «фагоцитоз» вживають тоді, коли клітина захоплює тверді частинки, а «піноцитоз» — коли вбираються краплини рідини. Розрізняють два види піноцитозу: ендоцитоз (відбувається вбирання рідини, проникнення її всередину клітини), екзоцитоз (відбувається виділення з клітини продуктів синтезу і життєдіяльності через плазматичну мембрану)’.
В одноклітинних організмів піноцитоз пов’язаний з процесами внутріклітинного травлення і є одним з важливих способів вбирання поживних речовин. Це так званий амебоїдний тип піноцитозу. Для іншого виду клітин, зокрема тих, функції яких пов’язані з процесами всмоктування, характерним є субмікроскопічний піноцитоз.
Ендоцитоз поділяється на кілька стадій і починається з адсорбції на поверхні плазматичної мембрани речовин, які вбираються. Потім відбувається вгинання мембрани з утворенням каналів, заповнених речовиною, яка вбирається, а краї мембрани на місці вгинання змикаються, в результаті чого утворюється ендоцитозний міхур, який мігрує в цитоплазму. Тут під впливом ферментів мембрана, яка оточує міхур, розчиняється і звільняє захоплену речовину.
Молекулярні механізми ендоцитозу з’ясовано не повністю. Вважають, що деякі етапи цього процесу потребують певних витрат: енергетичних і АТФ у вигляді джерела енергії.
Ендоцитоз речовин, які мають білкову природу, відбувається з участю специфічних рецепторів. У цьому випадку піноцитоз опосередкований рецепторами. Такий механізм потрапляння в клітину характерний для ряду гормонів (інсуліну), імуноглобулінів, ліпопротеїдів.
Екзоцитоз забезпечує транспорт і виділення з клітини речовин, які синтезуються — білків, гормонів, ферментів.
Як і ендоцитоз, він поділяється на кілька стадій. Спочатку синтезована речовина потрапляє на цистерни комплексу Гольджі, де відбувається її модифікація і «упаковка» в спеціальні секреторні екзоцитозні гранули, які з участю внутріклітинних скоротливих систем транспортуються до цитоплазматичної мембрани. Внаслідок безпосереднього контакту екзоцитозних гранул з цитоплазматичною мембраною відбувається їх злиття і перехід транспортованих речовин у позаклітинний простір.
Речовини з клітини можуть виділятись в результаті зворотного піноцитозу. При цьому захоплюються речовини плазматичною мембраною і відокремлюються утворені пухирці. Екзоцитоз особливо характерний для речовин ліпідної природи, які мають високу спорідненість з цитоплазматичними мембранами.
Явище фагоцитозу вперше було відкрито в кінці XIX ст. видатним російським біологом І. І. Мечниковим. Він встановив, що білі кров’яні тільця — лейкоцити мають здатність поглинати бактеріальні клітини. Згодом такі самі властивості було виявлено в інших клітин. Процес поглинання дістав назву фагоцитозу. При фагоцитозі бактерії чи інші тверді частинки захоплюються виростами цитоплазми і втягуються всередину клітини, де вони перетравлюються з участю гідролітичних ферментів. При цьому частинки поглинутого матеріалу можуть засвоюватись клітиною. Процес фагоцитозу відіграє важливу роль у захисних реакціях організму. Так, з участю спеціалізованих клітин лейкоцитів (фагоцитів) відбувається захоплення хвороботворних бактерій та їх знешкодження. Як бачимо, біологічні мембрани динамічні, оскільки в результаті фагоцитозу відбуваються всі види перетворень біомембран — перебудова, переміщення, злиття тощо.
Цитоплазма — основна складова частина внутріклітинного вмісту. Вона являє собою живу колоїдну систему (в’язкопружний тиксотропний гель) і має впорядковану субмікроскопічну структуру. До складу цитоплазми входять 75—85 % води, 10—12 % білків, 4—6 % вуглеводів, 2— 3% жирів та 1 % неорганічних речовин.
Завдяки наявності в’язкопружних властивостей цитоплазма одночасно має властивості в’язкої рідини і твердого тіла. Вона здатна до текучості, еластична. Як тиксотропний гель цитоплазма може змінювати ступінь в’язкості від рідкого стану (стан золю) до драглистого (стан гелю). Перехід з одного стану в інший відбувається під впливом різних внутріклітинних і зовнішніх факторів.
У багатьох клітинах зовнішній шар цитоплазми (ектоплазма), в якому міститься велика кількість органел, перебуває в стані гелю, а внутрішній (ендоплазма) тиксотропний. Вважають, що важливу роль у забезпеченні тиксотропності цитоплазми відіграють мікротрубочки і мікрофіламенти — тонкі видовжені нитчасті білкові мікроструктури немембранної природи. Мікротрубочки в клітині утворюють своєрідну сітку — цитоскелет. Найчастіше вони лежать в ектоплазмі паралельно плазматичній мембрані і забезпечують рух спеціалізованих клітин. Мікрофіламенти розміщуються пучками безпосередньо під цитоплазматичною мембраною і визначають внутріклітинний рух і текучість цитоплазми.
У більшості евкаріотичних клітин рух цитоплазми здійснюється з швидкістю 1—6 см/год і забезпечує оптимальне розміщення органоїдів, що сприяє протіканню біохімічних процесів і видаленню продуктів обміну, оскільки органели клітини теж рухаються разом з цитоплазмою. Рух цитоплазми у найпростіших забезпечує переміщення їх у просторі. Так, у амеб внаслідок скорочення актинових і міозинових мікрофіламентів відбувається локальне переміщення ендопла/зми з однієї частини клітини в іншу, що забезпечує рух клітини. Завдяки текучості цитоплазми відбувається транслокація хлоропластів залежно від освітлення, а також клітинних ядер, міхурців. У рослинних клітинах може здійснюватись і круговий потік цитоплазми навколо центральної вакуолі.
Гомогенна субстанція цитоплазми, розміщена між мікро-філаментами, називається матриксом. До складу матриксу входять вода і розчинені в ній мінеральні та органічні речовини, проміжні продукти обміну, ферменти, які забезпечують протікання у клітині таких досить важливих процесів обміну вуглеводів, як гліколіз і пентозофосфатний цикл.
Матрикс цитоплазми разом з деякими легкими структурами, які не осаджуються при ультрацентрифугуванні, має назву цитозолю, хоч в клітині матрикс міститься у вигляді в’язкоеластичного тиксотропного гелю.
У цитоплазмі рослинних і тваринних організмів містяться різноманітні органели. Вони є обов’язковими структурними компонентами внутріклітинного середовища і визначають перебіг багатьох метаболічних реакцій, без яких неможливо уявити функціонування живої системи. Усі вони мають певну форму, розміри, хімічний склад та виконують специфічні функції. Про важливість внутріклітинних
органел свідчить той факт, що після видалення їх клітина втрачає здатність нормально функціонувати і з часом гине.
Найважливіші органели клітини — ядро, мітохондрії, рибосоми, пластиди, вакуолі.
Ядро. Враховуючи функціональне значення ядра в забезпеченні метаболічних і генетичних процесів, його називають центром управління клітини або її інформаційним центром. Головні функції ядра полягають у збереженні і передачі інформації в цитоплазму з участю транскрипції та передачі генетичної інформації дочірнім клітинам внаслідок реплікації при поділі клітини.
Ще в 30-х роках минулого століття було сформульовано клітинну теорію, однак роль ядра як інформаційного центру клітини, в якому містяться хромосоми, що несуть спадкову інформацію в формі ДНК, було встановлено значно пізніше.
Найчастіше ядро розміщене в центрі клітини і може мати кулясту, овальну лінзоподібну, яйцеподібну форми, як і клітини, в яких воно міститься. Діаметр ядра в середньому становить 5 мкм. Ця величина може змінюватись від 0,5 мкм (у грибів) до 500 мкм (у яйцеклітинах). У рослинних клітинах, які мають центральну вакуолю, ядро розміщене в екзоплазмі і має сплющену, лінзоподібну форму.
Наявність в клітині відмежованого мембраною ядра є основною ознакою, за якою відрізняють евкаріоти від прокаріотів. Серед деяких евкаріотичних клітин зустрічаються без’ядерні клітини. Такі клітини (еритроцити, клітини ситоподібних трубок у вищих рослин) недовговічні і не здатні до поділу. Деякі клітини можуть мати кілька ядер. Такі багатоядерні клітини часто утворюються внаслідок злиття кількох клітин (поперечносмугасті м’язи, деякі судини рослий). Одноклітинні евкаріоти бувають двоядерні. Одне з ядер є джерелом генетичної інформації (мікронуклеус), а друге — здійснює керування метаболічними процесами (макронуклеус). Проте більшість клітин рослинних і тваринних організмів мають одне ядро.
Для кожного виду клітин існує певне співвідношення між розміром ядра і об’ємом цитоплазми. Незалежно від форми і кількості ядер у клітині всі вони складаються з нуклеоплазми, хромосом (хроматину), ядерець і ядерної оболонки. Ядерна оболонка має дві мембрани заввишки 6—8 нм, розділених перинуклеарним простором, ширина якого 10—40 нм. Зовнішня ядерна оболонка зв’язана з ендоплазматичним ретикулумом і утворюється наприкінці клітинного поділу внаслідок злиття цистерн ендоплазматичного ретикулуму з фрагментами старої ядерної оболонки, зруйнованої під час поділу.
В ядерних мембранах містяться пори (видимі в електронний мікроскоп), які забезпечують перехід з ядра в цитоплазму порівняно великих частинок. У більшості випадків пори мають восьмигранну форму і займають приблизно 5 % поверхні ядра. Вважають, що рибосомні білки, ферменти, гістони потрапляють в ядро з цитоплазми через пори, а дрібніші молекули та іони проникають з внутрішнього простору ендоплазматичного ретикулуму в перинуклеарний простір, а потім з участю системи активного транспорту проникають через внутрішню ядерну оболонку.
Всередині ядра міститься нуклеоплазма (каріолімфа), що складається з рідкої частини — ядерного матриксу і різних включень. Ядерний матрикс за хімічним складом подібний до матриксу цитоплазми і містить ферменти та проміжні продукти метаболізму. Серед включень найчастіше зустрічаються гранулярні або нитчасті рибонуклеопротеїдні частинки, ядерні тільця, що складаються з вуглеводів і ліпідів. В ядрі виявляється ряд структур — сітка з тонких ниток, між якими розміщена порівняно щільна забарвлена маса — хроматин, з якого під час мітозу утворюються хромосоми.
Хромосоми — це витягнуті в довжину нуклеопротеїдці частинки, які досить легко можна виділити як з прокаріотичних, так і з евкаріотичних клітин. Кількість, форма і розміри хромосом характерні для кожного виду рослин і тварин. Між певним вмістом хромосом (п) розділений весь фонд генетичної інформації клітинного ядра — геном. Ця кількість хромосом (п) специфічна для кожного виду — у кукурудзи 10, у людини — 23, у деяких водоростей — 600. Гаплоїдні клітини містять один набір хромосом, диплоїд- ні — 2, поліплоїдні — кілька.
Соматичні клітини вищих рослин і тварин диплоїдні і містять один батьківський і один материнський набір хромосом. Статеві клітини гаплоїдні. Гаплоїдні клітини утворюються з диплоїдних внаслідок мейозу, а диплоїдні — з гаплоїдних в результаті запліднення.
Хроматин прокаріотичних клітин складається лише з ДНК, а в евкаріотичних клітинах до його складу входять чотири компоненти: ДНК, РНК, основний низькомолекулярний білок-гістон і кислий негістоновий білок. ДНК в складі хроматину утворює з гістоном досить міцний нуклеопротеїдний комплекс — нуклеосому. У середньому до складу хроматину входять 40 % ДНК, 40 % гістонів, 20 % негістонових білків і незначна кількість РНК. Негістонові білки виконують досить різноманітні функції — здійснюють рух хромосом (актин і міозин), мають ферментні властивості (полімерази, кінази, метилази), регулюють активність певних генів. ДНК хромосом містить спадкову інформацію і забезпечує передачу її новоутвореним клітинам. Крім того, в ядрі здійснюються синтез іРНК, яка є матрицею для синтезу білкових молекул. Протягом клітинного циклу відбувається зміна двох фізіологічних форм хромосом — транспортної (під час поділу клітин) і функціональної (в інтер- фазі). Транспортні хромосоми компактні, паличкоподібні, а функціональні — розпушені, нитчасті. Розпушення хромосом необхідна умова транскрипції — синтезу іРНК. По всій довжині хромосоми проходить безперервна подвійна спіраль ДНК, яка у вищих організмів може містити 1-Ю8 пар азотистих основ. Уздовж молекули ДНК розміщені лінійні функціональні ділянки — гени, які становлять до 25 % її маси і несуть інформацію про синтез іРНК. Середня довжина гена 1000 пар основ.
Крім хроматину, в ядрі містяться округлі, досить ущільнені ділянки клітинного ядра — ядерця. Кількість їх може бути різною — від одного до семи. Ядерця прикріплені до певної ділянки однієї з хромосом, яка називається організатором ядерця. Внутрішня структура ядерця складається з рихлої сітки, що містить рибонуклеопротеїдні фібрили, гранулярні структури — рибонуклеопротеїдні гранули і основну масу, що складається з білків і РНК. До складу ядерець входять 80 % білків і 15 % РНК. Крім того, в них міститься незначна частина ядерцевого хроматину, обгорнутого рибонуклеопротеїдними фібрилами.
В ядерцях відбувається досить важливий процес — синтез рибосомної РНК. Під час мітозу ядерця зникають і синтез рибосомної РНК припиняється. Після закінчення поділу відбувається їх утворення на певних ділянках розпушеного хроматину.
Еквівалент ядра прокаріотичних клітин. У прокаріотичних одноклітинних організмах немає відокремленого ядра. Замість ядра вони містять його еквівалент — полінуклеотид, який не відмежовується різко від цитоплазми, що його оточує. Складається він з подвійної спіралі ДНК, закрученої і замкнутої в кільце, що прикріплена до плазматичної мембрани. ДНК прокаріотів, на відміну від ядерної ДНК евкаріотів, не зв’язана з гістонами чи іншими ядерними білками. Вона містить розміщені лінійно ді
лянки— гени, які забезпечують реплікацію. Так організований еквівалент ядра у бактерії кишкової палички, інші бактерії можуть містити ДНК у формі крихітних плазмід, що подібні до позаядерних ДНК евкаріотів.
Плазміди являють собою короткі подвійні спіралі ДНК, замкнуті в кільце, що перебувають у вільному стані, не включені в геном. До їх складу входять в середньому 100 000 пар основ і кілька генів, реплікація яких відбувається незалежно від основного генетичного матеріалу. Часто плазміди переходять з однієї клітини в іншу. Виявлено плазміди в прокаріотичних клітинах, а також у мітохондріях евкаріотичних клітин.
Рибосоми — обов’язкові самостійні клітинні органоїди. Вони мають вигляд грибоподібних тілець діаметром 0,015—0,02 мкм, що складаються з двох неоднакових за розміром частинок (мал. 4). Ці органели мають важливе значення для процесів білкового синтезу. Саме вони є безпосереднім місцем трансляції генетичної інформації. У процесі білкового синтезу окремі рибосоми об’єднуються, іноді відбувається об’єднання їх у групи з утворенням полісом. Структурну основу рибосом становлять специфічні рибосомальні РНК і білки. Найбільша кількість рибосом міститься в цитоплазмі клітин, в яких відбуваються активні білок- синтезуючі процеси — у клітинах, що ростуть, клітинах зародків, меристематичних клітинах. Крім того, рибосоми містяться в інших клітинних включеннях — хлоропластах і мітохондріях, де вони також беруть участь у синтезі білкових молекул.
Мітохондрії і пластиди є в більшості евкаріотичних клітин. Вони подібні за функціями, морфологічними особливостями і походженням. З їх участю в клітині відбуваються основні етапи перетворення енергії, тому їх називають енергетичними силовими підстанціями клітини. Водночас органели мають ряд особливостей, що свідчать про їх певну структурну і функціональну автономію. Вони містять здатну до реплікації ДНК, різні види РНК, рибосоми — свого роду білоксинтезуючий апарат, який дещо відрізняється від його ядерних і цитоплазматичних аналогів. За своїми властивостями він наближається до відповідного апарату прокаріотичних клітин — бактерій і синьо-зелених водоростей. Є дані, що частина білків і РНК цих органел синтезуються на їх власних рибосомах, тобто органели деякою мірою контролюють утворення частини своїх компонентів. Крім того, мітохондрії і пластиди виникають внаслідок поперечного поділу існуючих органел або розвиваються з маленьких міхурців, що мають подвійні мембрани та щільний матрикс і відокремлюються від зрілих мітохондрій чи пластид. У зв’язку з цим існує думка про симбіотичне походження цих органел. Вважають, що вони є потомками прокаріотів, подібних до бактерій і синьо-зелених водоростей, які проникли в клітину внаслідок фагоцитозу як «агресори», і лише в процесі еволюції створилась симбіотична асоціація з функціями, життєво важливими для клітини. За іншими гіпотезами мітохондрії виникли з вгинань цитоплазматичної мембрани, якими були оточені частини примітивного геному.
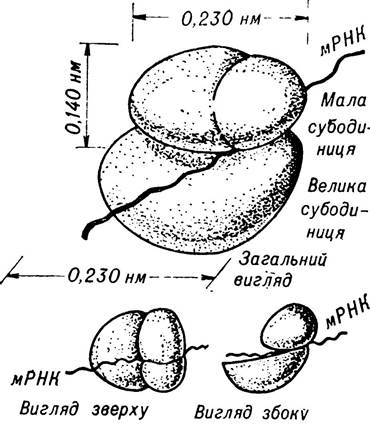
Мал. 4. Рибосоми печінки (за Nonomura, 1971).
Мітохондрії є складовими компонентами більшості клітин, крім клітин бактерій, синьо-зелених водоростей, зрілих еритроцитів крові, клітин деяких найпростіших, які ведуть паразитичний спосіб життя і здобувають енергію за рахунок процесів бродіння. У прокаріотів усі процеси енергозабезпечення відбуваються в плазматичній мембрані та її вгинаннях-тилакоїдах. Кількість мітохондрій і їх форма залежать від типу клітин, в яких вони містяться. У середньому в клітинах їх буває 150—1500, хоч у деяких найпростіших з інтенсивним обміном їх може бути до 500 000.
У рослинних клітинах мітохондрій значно менше, ніж у тваринних. Здебільшого вони мають форму паличок, кульок, лінз, ниток.
На поверхні мітохондрій міститься подвійна мембрана, між її частинами є перимітохондріальний простір. Значна частина ферментів, що перебуває в цьому просторі, забезпечує з участю АТФ процеси фосфорилювання різних субстратів. Товщина мембран 7—10 нм. Зовнішня мембрана має гладку поверхню, а внутрішня утворює велику кількість виступів, складок, перегородок, трубочок, що називаються кристами. Кристи пронизують внутрішній простір мітохондрій, де вони утворюють своєрідні відсіки (камери), внаслідок чого значно збільшується поверхня внутрішньої мембрани. Між кристами міститься основна речовина — матрикс, за складом подібний до цитоплазми.
Співвідношення між матриксом і кристами в різних мітохондріях залежить від їх функцій і ступеня біологічної активності. У мітохондріях, в яких відбуваються біосинтетичні процеси, переважає матрикс і дуже мало крист (мітохондрії печінки), а в мітохондріях, які спеціалізуються на виробництві енергії, крист більше. У матриксі містяться гранули, до складу яких входять білки, фосфоліпіди, а також різні іони (Са2+, Mg2+) та фосфати, ДНК, різні види РНК, рибосоми. У мітохондріальній ДНК закодовані мітохондріальні транспортні і рибосомні РНК, деякі білки внутрішньої мембрани мітохондрій — цитохром b, субодиниці цитохромоксидази.
Зовнішні і внутрішні мембрани мітохондрій різняться між собою хімічним складом і мають деякі свої особливості. Зовнішні мембрани містять велику кількість складних ліпідів і холестерину. Хімічний склад їх подібний до плазматичної мембрани. Через численні пори зовнішньої мембрани, утворені тунельними білками, досить легко проходять порівняно великі молекули — амінокислоти, цукри, АТФ. На зовнішній мембрані мітохондрій локалізовані ферменти обміну фосфоліпідів, активації жирних кислот і моноамінооксидаза. Внутрішні мембрани мітохондрій містять до 75 % білків і незначну кількість ліпідів, серед яких переважає кардіоліпін. На відміну від зовнішніх, внутрішні мембрани практично непроникні для більшості метаболітів, що містяться в матриксі,— глюкози, пірувату, амінокислот, АТФ, АДФ, фосфатів. Тому транспорт цих речовин відбувається з участю специфічних транспортних білків і з використанням енергії АТФ (активний транспорт).
У внутрішній мембрані мітохондрій і кристах містяться ферментні системи, які забезпечують протікання численних реакцій, пов’язаних з процесами утворення і нагромадження енергії — перетворення хімічних зв’язків поживних речовин в енергоємні зв’язки АТФ. У вигляді інтегральних білків тут міститься комплекс ферментів, які беруть участь у транспорті електронів (дихальний ланцюг). До складу периферичних білків входять різні дегідрогенази, які беруть участь в окисленні субстратів дихання і передачі водн., знятого із субстрату, на дихальний ланцюг.
За допомогою електронного мікроскопа на внутрішніх мембранах мітохондрій і крист можна спостерігати грибоподібні утворення— мембранні АТФ-ази, які здійснюють синтез АТФ, а також зв’язуючі фактори, що забезпечують енергетичний зв’язок між процесами дихання і окислювального фосфорилювання. Енергія, акумульована в макроергічних зв’язках АТФ, використовується для забезпечення різноманітних метаболічних процесів — росту, розвитку, синтезу.
Пластиди характерні лише для рослинних клітин і є найбільшими (після ядра) цитоплазматичними внутріклітинними структурами. Усі вони розвиваються з одного спільного типу безбарвних пропластид, які далі залежно від виду тканин, в яких вони функціонують у зрілому вигляді, розвиваються в зелені хлоропласти або у хромопласти та лейкопласти. Хлоропласти — найпоширеніший тип пластид рослинних клітин, основні органоїди, в яких відбувається складний процес перетворення світлової енергії в хімічну енергію органічних сполук. Цей унікальний, не до кінця розгаданий процес, називається фотосинтезом. У хлоропластах міститься 50—70 % води, решта — суха речовина, що складається з 36—50 % білків, 29—30 % ліпідів, 6—8 % мінеральних речовин, 1 % хлорофілу, 8—10 % вуглеводів та інших речовин — вітамінів, нуклеїнових кислот, пігментів.
Значна кількість хлоропластів міститься в клітинах тканин, які утворюються на світлі,— в листках, стеблах, молодих плодах. На площу 1 мм2 листкової пластинки припадає приблизно 400 000 хлоропластів. У клітинах водоростей міститься переважно 2—3 хлоропласти, а в клітинах вищих рослин їх значно більше — 20—50. Хлоропласти водоростей мають вигляд сітки, зірчастої пластинки чи спіралеподібної стрічки.
Будова хлоропластів вищих рослин значно складніша. Подібно до мітохондрій тваринних клітин вони мають зовнішню і внутрішню мембрани, які оточують безбарвну основну речовину — матрикс або строму.
Із складом внутрішньої мембрани всередині хлоропластів утворюється розгалужена система двомембранних структур — ламел. Кінці ламел попарно з’єднані так, що кожні дві сусідні утворюють порожнисті мембранні кармани, локальне скупчення яких називається тилакоїдами. По- парне з’єднання ламел дає змогу розрізняти в них внутрішню й зовнішню поверхню. Тилакоїди групуються в щільно упаковані диски або грани величиною 0,5—0,8 мкм. У хлоропластах буває 10—100 гран. Вони з’єднуються між собою міжгранними ламелами, які пронизують строму хлоропластів. Ламели гран мають товщину 4—6 нм, міжгранні ламели — 2—3 нм. Таким чином, всередині хлоропластів створюється ламелярно-гранулярна структура.
Для всіх вищих рослин принцип будови хлоропластів однаковий, хоч хлоропласти різних видів можуть різнитись між собою величиною та кількістю ламел. Мембрани ламел містять до 50 % білків. Ліпіди відіграють важливу роль у формуванні мембран та у створенні їх гідрофобних діелектричних шарів. Білки пронизують товщу мембран і одночасно з зеленим пігментом хлорофілом виробляють сфероїдні частинки — фотосистеми, в яких відбуваються важливі реакції, пов’язані з вбиранням світлової енергії.
У стромі хлоропластів міститься замкнута в кільце молекула ДНК, яка несе генетичний матеріал. Молекули ДНК хлоропластів, як і мітохондрій, не асоційовані з гістоновими білками, однак ДНК хлоропластів більша за розмірами і несе більше інформації. Вона кодує різні види РНК (тРНК, РНК-полімерази), деякі види білків рибосом, пластидні цитохроми та значну кількість ферментів певних фаз фотосинтезу.
Лейкопласти — безбарвні пластиди округлої, овальної або веретеноподібної форми, розміщені в цитоплазмі неосвітлених частин рослин. Найбільша кількість їх міститься в клітинах кореневищ, насіння, епідермісу, бульбах, серцевині стебел. Лейкопласти значно відрізняються від хлоропластів як за розмірами (вони значно менші), так і за внутрішньою будовою. У лейкопластах майже немає тилакоїдів — вгинань і складок внутрішньої мембрани. Утворення їх або генетично загальмоване, як у клітинах коріння та епідермісу, або ж гальмується відсутністю світла (у клітинах бульб). В останньому випадку від надмірного освітлення може перебудовуватись внутрішня структура лейкопластів і перетворюватись у хлоропласти. Саме тому внаслідок тривалого освітлення бульби картоплі часто зеленіють. Крім поодиноких тилакоїдів, всередині лейкопластів містяться крохмальні зерна, ДНК, пластоглобули і пластидний центр (проламелярне тільце).
У крохмальних зернах відкладаються запасні вуглеводи у вигляді крохмалю, який синтезується з участю відповідної ферментної системи з надлишків глюкози, утвореної в. результаті фотосинтезу. Крохмальні зерна різних рослин різняться між собою формою і внутрішньою структурою, тому є важливою таксономічною ознакою.
Пластидний центр лейкопластів складається з переплетеної сітки розгалужених трубочок або різних тілець і міхурдів. Саме з пластидного центру на світлі можуть формуватись тилакоїди.
Хромопласті! визначають гаму забарвлень плодів, квіток та інших частин рослини. Утворюються вони з хлоропластів або лейкопластів. При цьому руйнуються хлорофіл і тилакоїди та вивільнюються каротиноїди або відбувається синтез їх. Це явище спостерігається під час дозрівання фруктів і овочів, зміни забарвлення листя дерев восени. У пелюстках деяких квіток (губоцвітих) хромопласти є самостійними органелами.
Хромопласти містять переважно жовті пігменти — каротиноїди, яких налічується 50 видів. Відкладаються ці пігменти в пластоглобулах — трубчастих або ниткоподібних білкових структурах. Внутрішня структура хромопластів виражена слабо. Крім пластоглобул, у них містяться крохмальні зерна й білкові кристалоїди. Тилакоїдів у них майже немає, як немає й пластидного центру. Тому для хромопластів не властива функціональна активність. їх роль полягає в забезпеченні забарвлення певних частин рослини.
У клітинах існують різні структури й функціональні модифікації пластид, однак лише хлорофіловмісні пластиди з тилакоїдним (ламелярно-гранулярним) типом організації можуть забезпечувати перетворення світлової енергії в енергію хімічних зв’язків органічних сполук.
Крім плазматичної мембрани, у клітині існує ціла система ендомембран, таких, як ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, мікротільця і вакуолі. Усі вони утворюють обмежені мембранами відсіки клітини, що характеризуються певними функціональними особливостями.
Ендоплазматичний ретикулум. В усіх евкаріотичних клітинах міститься внутрішньоплазматична сітка, що складається з досить розгалужених і переплетених мембран. Мембрани ендоплазматичного ретикулуму утворюють систему каналів, трубочок, цистерн різних розмірів і форм, які часто пов’язані між собою, а їх внутрішній простір межує з перинуклеарною частиною клітини. Різні клітини характеризуються не однаковим ступенем розвитку ендоплазматичного ретикулуму та його компонентів, що визначається метаболічною активністю клітини. У рослинних клітин трубчасті цистерни можуть проникати через клітинну стінку в сусідні клітини.
Внаслідок виділення ендоплазматичного ретикулуму під час гомогенізації він руйнується, утворюючи фрагменти певного розміру — мікросоми. Мембрани цистерн ендоплазматичного ретикулуму містять велику кількість гліцерофосфатидів, вміст яких може становити 95 %. Товщина мембран 6 нм.
Розрізняють два типи ендоплазматичного ретикулуму залежно від наявності чи відсутності на його поверхні специфічних органел клітини — рибосом. У разі відсутності рибосом утворюється ендоплазматичний ретикулум з гладенькими стінками — агранулярний ретикулум, а якщо на його поверхні є невеликі гранули (рибосоми)—ендоплазматичний гранулярний ретикулум.
Гранулярний ендоплазматичний ретикулум добре розвинений у клітинах, де відбувається інтенсивний синтез білків. Рибосоми часто об’єднуються в спіралеподібні утворення — полісоми, що прикріплюються до зовнішньої поверхні мембран ендоплазматичного ретикулуму своїми більшими субодиницями і беруть безпосередню участь у процесах білкового синтезу. Синтезовані білки з рибосом проходять через мембрану в цистерни ендоплазматичного ретикулуму, звідки через трубчасті елементи ретикулуму виводяться в позаклітинний простір. Таким чином, ендоплазматичний ретикулум забезпечує синтез і транспорт важливих біополі- мерів. З його участю створюється транспортна система клітини, яка розділяє синтезовані продукти і переносить їх у різні ділянки клітини чи в позаклітинний простір.
Агранулярний ендоплазматичний ретикулум утворює трубчасті елементи — канальці різної довжини. Діаметр порожнини трубочок становить 0,025—0,03 мкм. Завдяки конструкції з порожнистих пористих трубочок створюється поверхня, на якій відбувається послідовно чи одночасно перебіг різноманітних хімічних реакцій.
Трубчасті пористі елементи гладенького ендоплазматичного ретикулуму беруть участь у синтезі й транспорті різних речовин як всередині клітини, так і поза нею. Вони також забезпечують різні етапи метаболізму вуглеводів, ліпідів, стероїдів. Особливо інтенсивно розвинений агранулярний ендоплазматичний ретикулум у клітинах, в яких синтезуються ліпіди і мембранні стероїди (холестерин). Значна кількість його міститься в клітинах сім’яників і надниркових залоз, де синтезуються стероїдні гормони. З мембранами агранулярного ендоплазматичного ретикулуму зв’язані ферменти, які допомагають забезпечити обмін вуглеводів, ліпідів, а також ферментні системи дихального ланцюга.
У клітині між елементами гранулярного і агранулярного ретикулуму існує тісний взаємозв’язок. Вони є частиною однієї й тієї самої лабільної мембранної системи, яка має здатність до внутріклітинного диференціювання і певних видозмін.
Система Гольджі. Майже в усіх евкаріотичних клітинах міститься система мембран, подібна до ендоплазматичного ретикулуму, яка дістала назву від прізвища вченого, що перший описав її. Система Гольджі складається, як правило, з кількох відособлених комплексів Гольджі, або диктіосом. До складу диктіосоми входять 3—12 дископодібних замкнутих цистерн Гольджі, що розміщені паралельно і покриті оболонкою. Цистерни Гольджі досить часто бувають вигнуті, що створює випуклу зовнішню і ввігнуту внутрішню поверхню. Кількість цистерн у клітинах різна, що залежить від їх виду. Часто між цистернами переплітаються тонкі трубчасті або фібрилярні елементи, які розгалужуються в різні боки від клітини. Внаслідок злиття всіх диктіосом утворюється зернисто-зірчаста структура, яка іноді розміщується у вигляді ковпачка безпосередньо в ядерній зоні клітини, створюючи систему або апарат Гольджі. Від зрілих диктіосом поступово відокремлюються дрібні міхурці, заповнені секретом, і мігрують до периферії системи. Крім міхурців, до складу апарату Гольджі входять також об’ємні вакуолі, які утворюються внаслідок розширення цистерн. Сформовані міхурці і вакуолі підходять до плазматичної мембрани, зливаються з нею, в результаті чого здійснюється перехід секрету в позаклітинний простір (екзоцитоз). Інколи міхурці можуть зливатися також із секреторними гранулами всередині клітини. В зв’язку з цим основною функцією апарату Гольджі є нагромадження, упаковка і транспорт за допомогою міхурців продуктів внутріклітинного метаболізму — синтезованих речовин, продуктів розпаду і токсинів.
Вважають, що система Гольджі є похідним ендоплазматичного ретикулуму. Злиття окремих частин ендоплазматичного ретикулуму (міхурців і фрагментів) веде до формування нових цистерн Гольджі. Одночасно з цим відбувається перебудова мембран ендоплазматичного ретикулуму в щільнішу мембрану апарату Гольджі з іншим ліпідним і білковим складом, тобто між цими двома мембранними системами існує тісний взаємозв’язок.
Вакуолі характерні для зрілих рослинних клітин, вони являють собою міхурці, заповнені водянистим вмістом — клітинним соком і оточені елементарною мембраною — тонопластом. Вони утворюються з ендоплазматичного ретикулуму або міхурців Гольджі.
Вакуолі відіграють важливу роль в регуляції тургору рослинних клітин, беруть участь у процесах росту, скорочення, нагромадження і збереження кінцевих продуктів обміну цих клітин.
Залежно від функцій, які вони виконують у клітинах, розрізняють кілька видів вакуоль. Для прісноводних найпростіших характерними є скоротливі (пульсуючі) вакуолі, що забезпечують їх осмотичну регуляцію. З участю пульсуючих вакуолей відбувається періодичне виділення з клітини надмірної кількості води, яка потрапляє з гіпотонічного навколишнього розчину чи вбирається на основі піноцитозу. Скорочуються вакуолі з участю еластичних волокон, які містяться в мембрані вакуолей.
У клітинах запасних тканин рослинних організмів часто зустрічаються вакуолі, в яких нагромаджуються поживні речовини — жири, білки тощо.
Досить розповсюдженою в рослинних клітинах, особливо в меристематичних, є центральна вакуоля, яка утворюється внаслідок злиття дрібних вакуолей. Центральна вакуоля часто займає 90 % внутріклітинного простору, відтісняючи компоненти цитоплазми і ядро до клітинної стінки, внаслідок чого підвищується інтенсивність обміну між цитоплазмою і зовнішнім середовищем.
Всередині вакуолі міститься клітинний сік. Мембрана вакуолі за властивостями нагадує мембрани ендоплазматичного ретикулуму, проте вона тонша, менш щільна і має більшу проникність. Оскільки внутрішній вміст вакуолі є гіпертонічним відносно цитоплазми і навколишнього середовища, то у вакуолі досить інтенсивно нагнітається вода, створюючи тургорний тиск. Тургорний тиск розтягує еластичну клітинну стінку і підтримує певні розміри клітин, їх жорсткість, що особливо важливо для молодих частин рослинних організмів, що ростуть.
Центральна вакуоля виконує в рослинних клітинах важливі функції і використовується для різноманітних цілей. Тут нагромаджуються і відособлюються проміжні продукти обміну — цукри, органічні кислоти, амінокислоти. У вакуолі нагромаджуються також різноманітні кінцеві продукти обміну, зокрема різноманітні пігменти, які зумовлюють забарвлення квіток, плодів (антоціани, флавони), а також токсичних продуктів — алкалоїдів, поліфенолів. Крім того, вакуолі можуть виконувати роль лізосомного простору, оскільки в них міститься певна кількість лізосомних ферментів, які потрапляють в них з міхурців Гольджі.
Лізосоми — це дрібні сферичні утворення, цитоплазматичні гранули розміром 0,2—0,5 мкм, оточені елементарною мембраною. Вони містять гідролітичні ферменти і забезпечують в клітині перетравлювання різних речовин. Всього в складі лісозом міститься біля ЗО різних ферментів.
Для лізосом характерним є явище гетеро- та аутофагії. Гетерофагія — де перетравлювання з участю гетеролізосом (фаголізосом) речовин, які потрапляють у клітину внаслідок ендоцитозу і фагоцитозу. Аутофагія — розщеплення в аутолізосомах (цитолізосомах) запасних внутріклітинних речовин, а також макромолекул чи органел, які втратили функціональну активність, зокрема мітохондрій. Крім того, внаслідок пошкодження лізосомних мембран, старіння чи патологічних змін ферменти потрапляють всередину клітини і починають перетравлювати внутріклітинний вміст. Відбувається автоліз, або лізис (самоперетравлювання протопласту), і клітина гине.
Лізосоми утворюються з цистерн Гольджі або агранулярного ендоплазматичного ретикулуму. Утворені первинні лізосоми являють собою гладенькі міхурці, що містять набір гідролітичних ферментів, синтезованих на гранулярному ендоплазматичному ретикулумі. Після вбирання субстрату первинні лізосоми перетворюються в функціонально активні вторинні лізосоми. Субстрат може вбиратись внаслідок аутофагії — активного вбирання макромолекул з цитоплазми, що його оточує. Крім того, лізосоми можуть зливатися з ендоцитезними чи фагоцитозними міхурцями (вакуолями) .
Іноді багато дрібних лізосом зливається, утворюючи велику за розміром первинну лізосому, яка може повністю захоплювати різні органели чи піноцитозні міхурці. Такі утворені вторинні лізосоми називаються мультивезикулярними тільцями.
Після закінчення процесу розщеплення ввібраних речовин вторинні лізосоми перетворюються в залишкові тільця, які містять неперетравлені залишки і зовсім не мають ферментів. Залишкові тільця або виводяться з клітини екзоцитозом, або розчиняються в цитоплазмі. Таким чином, лізосоми можна розглядати як систему видалення з клітини різноманітних продуктів обміну, що потрапляють з навколишнього середовища, а також видалення сторонніх тіл і структурних елементів клітини, які втратили певне функціональне значення.
Мікротільця, подібно до лізосом, оточені мембраною і містять певні ферменти. Це дрібні маложивучі міхурці (0,1—1,5 мкм) з гладенькими стінками і тонкозернистим
матриксом, який містить аморфні кристалоїдні включення. Основним компонентом матриксу мікротілець є білок. Головна функція цих органел полягає в закріпленні на певних ділянках клітини ферментів, які каталізують певні біохімічні реакції і таким чином забезпечують їх одночасне протікання в клітині. Вважають, що мікротільця виникають з розширених, заповнених ферментом цистерн ендоплазматичного ретикулуму.
Отже, розглянуті мембранні системи становлять обмежені мембранні відсіки клітини, що спеціалізуються залежно від функцій, які вони виконують. Між мембранами ендоплазматичного ретикулуму та іншими мембранними системами в клітині існує тісний взаємозв’язок. Вважають, що мембрани різних клітинних компонентів утворюються з мембран ендоплазматичного ретикулуму. Незважаючи на різноманітність мембранної системи, для всіх мембран, що її утворюють, характерним є спільний план будови, хоч вміст певних складових компонентів може бути різним.