БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 9.НУКЛЕЇНОВІ КИСЛОТИ
9.3. Фізико-хімічні властивості азотистих основ
Для азотистих основ характерним є існування таутомерних форм - тип ізомерії, за якої ізомери можуть переходити один у другий (рис. 9.1).
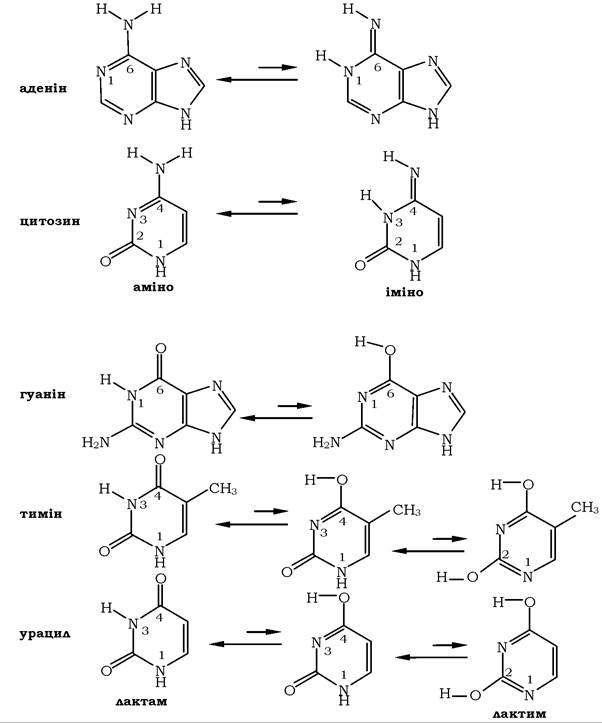
Рис. 9.1. Таутомери азотистих основ
В основі таутомерії лежить міграція протона, тобто внутрішньомолекулярні переміщення ядра водню від одного атома до іншого. Подібна міграція супроводжується зазвичай змінами електронної структури молекули. Азотисті основи, котрі входять до складу нуклеїнових кислот, залежно від структури гетероциклу здатні до аміно-імінної та кето-енольної (лактам-лактимної) таутомерії.
У складі нуклеїнових кислот аденін і цитозин перебувають переважно в аміноформі, а гуанін, тимін та урацил - у лактамній формі.
Азотисті основи слабко розчинні у воді. При нейтральному значенні рН найменшою розчинністю характеризується гуанін.
Завдяки гетероциклічній ароматичній природі пуринові й піримідинові азотисті основи поглинають електромагнітну енергію в ультрафіолетовому діапазоні (200-300 нм) і максимум поглинання становить близько 260 нм. Цю властивість використовують при кількісному визначенні нуклеїнових кислот.
Нуклеозиди - це N- або С-глікозидні похідні азотистих основ пуринового або піримідинового ряду. До складу нуклеозидів входять рибоза або дезоксирибоза, які містяться у фуранозній формі й завжди є D-ізомерами та β-аномерами.
Нумерація атомів у вуглеводному залишку залишається такою самою, як і у вільних пентоз, тобто починається від глікозидного центра. Але щоб відрізняти їх від номерів атомів азотистих основ, їх додатково позначають штрихами: атом вуглецю, зв'язаний з гетероциклом, нумерують як 1'-, атоми вуглецю з гідроксигрупами в рибонуклеотидів - 2'-, 3'-, 5'-, а компоненти ДНК - дезоксирибонуклеозиди - містять гідроксигрупи у 3'- та 5'-положеннях.
Вуглеводний залишок і азотиста основа зв'язані відносно кислотолабільним N-глікозидним зв'язком між першим атомом вуглецю пентози й першим атомом азоту піримідину або дев'ятим атомом азоту пурину. Залежно від типу пентози розрізняють два види нуклеозидів - рибонуклеозиди (містять рибозу та входять до РНК) і дезоксирибонуклеозиди (містять дезоксирибозу та входять до ДНК). Як приклад наведено структуру двох нуклеозидів:

Теоретично, залишок пентози та азотисті основи в нуклеозидах спроможні вільно обертатися навколо осі глікозидного зв'язку, проте насправді існують стеричні перешкоди цьому. Необхідною умовою для комплементарної взаємодії пуринових і піримідинових основ у дволанцюговій молекулі ДНК є анти- конформація молекул, і є оптимальнішою для природніх нуклеозидів ніж синконформація (рис. 9.2).
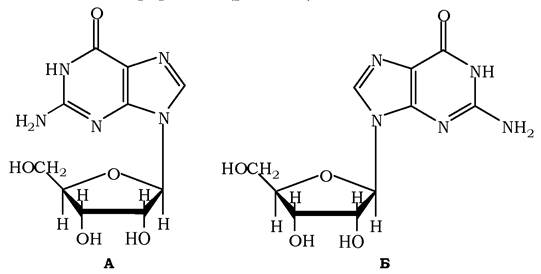
Рис. 9.2. Структура син- (А) та анти- (Б) конфігурацій гуанозину
Нуклеозиди не мають відновних властивостей, про що свідчить відсутність вільної альдегідної групи у вуглеводному компоненті. Нуклеозиди значно краще розчинні у воді, ніж вихідні азотисті основи. Крім цього, вони досить стійкі в нейтральних та лужних середовищах і гідролізуються за наявності мінеральних та органічних кислот. Швидкість гідролізу залежить від концентрації іонів водню. Пуринові нуклеозиди менш стійкі до гідролізу, ніж піримідинові.
Нуклеотиди - це фосфорнокислі ефіри нуклеозидів, у яких фосфорна кислота з'єднана складноефірним зв'язком з однією із вільних гідроксильних груп пентози. Усі нуклеотиди є сильними кислотами, оскільки залишок фосфорної кислоти легко дисоціює. Мономерні одиниці РНК, в яких вуглеводневою складовою є D-рибоза, називаються рибонуклеотидами, а мономерні одиниці ДНК, що включають D-дезоксирибозу - дезоксирибонуклеотидами. У рибонуклеотидах фосфат може приєднуватися в положеннях 2', 3', 5' D-рибози, а в дезоксирибонуклеотидах - у положеннях 3',5' D-дезоксирибози. Разом із тим унаслідок ферментативних реакцій синтезу чи розкладу нуклеїнових кислот усі клітини містять у вільній формі тільки нуклеозид-5-фосфати.
Фосфорилювання нуклеозидмонофосфатів (НМФ) у положенні С-5' атома вуглецю пентози приводить до утворення нуклеозиддифосфа- тів (НДФ) і нуклеозидтрифосфатів (НТФ). Як приклад наведено структури АТФ, а також відповідних ди- та монофосфатів (рис. 9.3).
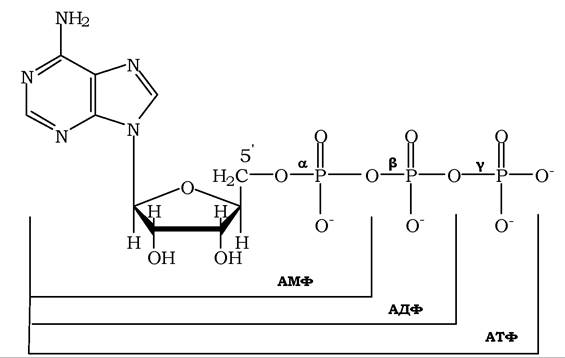
Рис. 9.3. Структура аденозинмоно-, ди- та трифосфатів
Усі нуклеозидфосфати містяться в клітині у вигляді аніонів, тому аденозинфосфати правильніше позначати АМФ2-, АДФ3-, АТФ4-. Назву нуклеотиди найчастіше отримують згідно з назвою відповідних нуклеозидів із зазначенням місця приєднання ортофосфату до залишку рибози або дезоксирибози. Поряд із цим використовують дещо вкороченні або скороченні назви. У табл. 9.2 наведено номенклатуру нуклеозидів і нуклеотидів.
Таблиця 9.2
Номенклатура азотистих основ, нуклеозидів і нуклеотидів
Азотисті основи |
||||
Назва |
Пурини |
Піримідини |
||
Аденін (А) |
Гуанін (Г) |
Цитозин (Ц) |
Урацил (У) Тимін (Т) |
|
Нуклеозиди: |
||||
РНК |
Аденозин |
Гуанозин |
Цитидин |
Уридин |
ДНК |
Дезоксі- |
Дезокси- |
Дезокси- |
Дезокси- |
аденозин |
гуанозин |
цитидин |
тимідин |
|
Нуклеотиди: |
||||
РНК |
Аденілат |
Гуанілат |
Цитидилат |
Уридилат |
ДНК |
Дезоксі- аденілат |
Дезокси- гуанілат |
Дезокси- цитидилат |
Тимідилат |
Нуклеозидмоно- |
АМФ |
ГМФ |
ЦМФ |
УМФ |
фосфати |
(дАМФ) |
(дГМФ) |
(дЦМФ) |
(ТМФ) |
Нуклеозидди- |
АДФ |
ГДФ |
ЦДФ |
УДФ |
фосфати |
(дАДФ) |
(дГДФ) |
(дЦДФ) |
(ТДФ) |
Нуклеозидтри- |
АТФ |
ГТФ |
ЦТФ |
УТФ |
фосфати |
(дАТФ) |
(дГТФ) |
(дЦТФ) |
(ТТФ) |
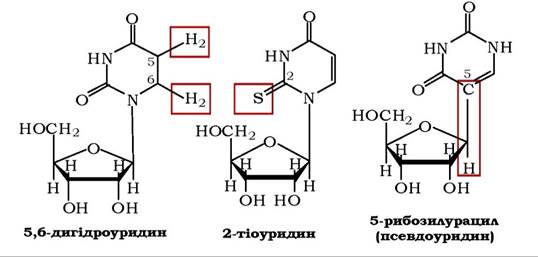
Поряд із головними компонентами, які входять до складу нуклеїнових кислот, інколи зустрічаються мінорні нуклеотиди (лат. minor - менший). Особливо різноманітні вони в тРНК. Мінорні нуклеотиди утворюються в організмі внаслідок хімічних перетворень (ковалентних модифікацій) нуклеотидних залишків, які відбуваються вже після формування полінуклеотидного ланцюга. Наприклад, залишки уридину в певних ділянках попередника РНК можуть піддаватися гідруванню, що приводить до утворення залишку дигідроуридину, сульфгідруванню з утворенням залишку тіоуридину, ізомеризації, яка зумовлює утворення залишку псевдоуридину (Ψ), де рибоза приєднана до урацилу в п'ятому положенні С-глікозидним зв'язком:
Надзвичайно поширені в РНК (а також у ДНК) метильовані похідні азотистих основ, до того ж у РНК об'єктом метилювання можуть бути і 2'-ОН групи залишків рибози. Нижче наведено структури різноманітних мінорних нуклеотидів, які зустрічаються в молекулах нуклеїнових кислот:
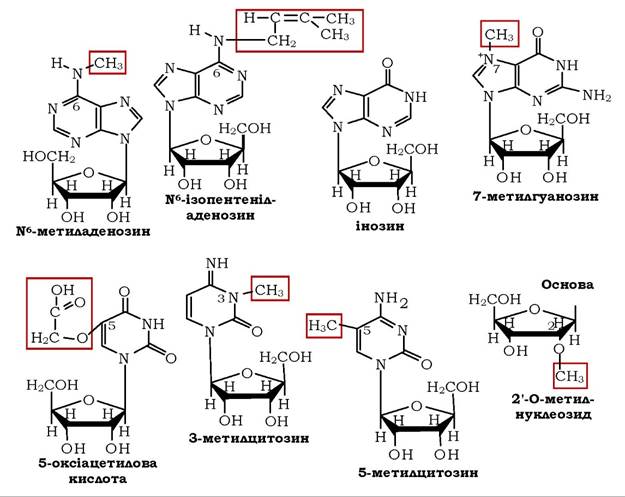
До похідних нуклеотидів, зокрема аденозинів, відносять активну форму амінокислоти метіоніну - S-аденозилметіонін, в якому сульфонієва структура з тризаміщеним атомом сірки (=S+-СH3) нестабільна й зумовлює високу активність метильної групи. Як донор метильних груп у реакціях метилювання біомолекул S-аденозилметіонін бере участь у синтезі креатину, в утворенні холіну з аміноспирту етаноламіну, адреналіну з норадреналіну, метилюванні азотистих основ нуклеотидів тощо.

Дезоксирибонуклеотиди в організмі використовуються виключно для біосинтезу ДНК. Функції рибонуклеотидів різноманітні:
✵ вони є попередниками для біосинтезу РНК;
✵ усі нуклеозидтрифосфати є макроергічними сполуками, хімічна енергія яких використовується організмом для синтезу біологічних речовин; при цьому унікальну роль у перетворенні енергії виконує АТФ, наприклад:
амінокислота + тРНК + АТФ → аміноацил-тРНК + АМФ + ФФн; жирна кислота + КоА-SH + АТФ → ацил-КоА + АМФ + ФФн; нікотинамідмононуклеотид + АТФ → НАД+ ФФн; нікотинамідаденіндинуклеотид + АТФ → НАДФ + АДФ;
✵ похідні нуклеотидів є донорами активних субстратів у синтезі гомо- та гетерополісахаридів, ліпідів і білків. Так, ГДФ- маноза бере участь у синтезі глікогену та глікозамінгліканів, ЦДФ-холін - у синтезі фосфоліпідів;
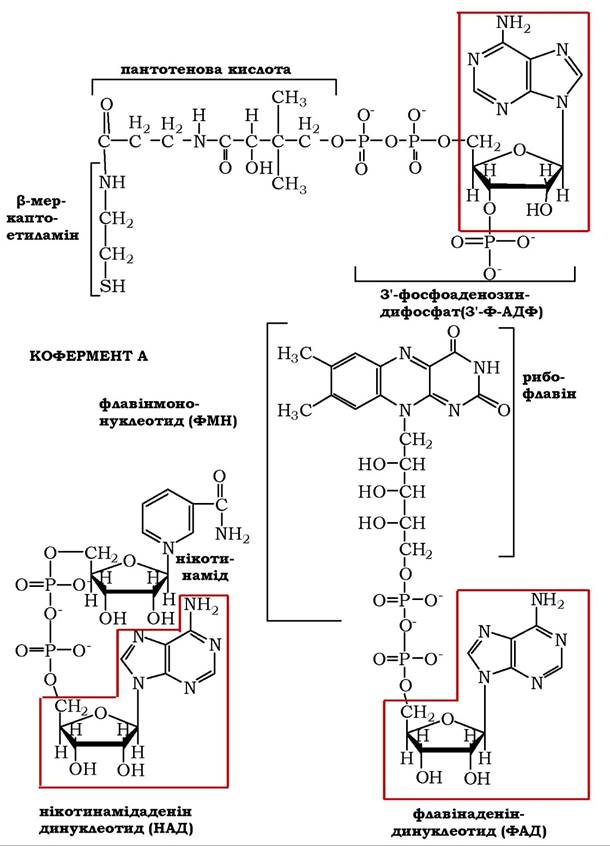
Рис. 9.4. Коферменти, у структурі яких бере участь аденозин
✵ S-аденозилметіонін (SAM), 3-фосфоаденозин-5-фосфосуль- фат (ФАФС), УДФ-глюкуронова кислота беруть участь в універсальній системі детоксикації, яка забезпечує виведення чужорідних речовин (ксенобіотиків) і деяких ендогенних метаболітів з організму;
✵ аденілові рибонуклеотиди (АМФ) входять до складу коферментів НАД, НАДФ, ФАД (ФМН) і коферменту ацилювання (КоА), які беруть участь у багатьох ферментативних реакціях. Структура таких коферментів зображено на рис. 9.4.
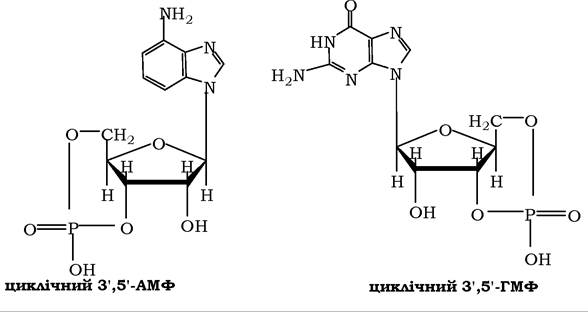
✵ окрему групу становить система циклічних нуклеотидів, у яких фосфатний залишок утворює складноефірні зв'язки з 3'- і 5'-гідроксильними групами рибози. В організмі вони відіграють роль внутрішньоклітинних (вторинних) посередників при передачі сигналів від гормонів, факторів росту, нейромедіаторів або інших позаклітинних регуляторів: