БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 10. МЕТАБОЛІЗМ НУКЛЕЇНОВИХ КИСЛОТ
10.7.Біосинтез РНК (транскрипція)
10.7.2.Механізм транскрипції ДНК
У процесі транскрипції розрізняють три стадії: ініціацію, елонгацію та термінацію.
Ініціація транскрипції відбувається внаслідок приєднання ДНК-залежної РНК-полімерази до матриці, але не на будь-якому місці, а в спеціальних ділянках, які називаються промоторними. Промотор (англ. promoter - той, хто сприяє чомусь) - стартова точка транскрипції, де відбувається впізнавання певних нуклеоти- дів ферментом. Бактеріальна РНК-полімераза безпосередньо впізнає одну з послідовностей у складі промотору на відстані 35 нуклеотидних пар до сайту ініціації (35-послідовність), яка складається з 8-10 нуклеотидів і є, як вважають, ділянкою, з якою взаємодіє σ-субодиниця холоферменту РНК-полімерази (рис. 10.22).
На ближчій відстані до сайту ініціації транскрипції - приблизно 10 основ (10-послідовність) до того нуклеотиду, з якого починається транскрипція, розташована шестичленна АТ-збагачена ділянка, яка має назву ТАТА-послідовність (бокс Прибнова). У даному сайті полімераза зв'язується з ДНК за допомогою спеціального білка - ТАТА-фактора. Приєднання ТАТА-фактора сприяє
взаємодії промотору з РНК-полімеразою і забезпечує локальне розходження приблизно 1,5 витка (15 нуклеотидних пар) спіралі ДНК. Завдяки дисоціації ланцюгів ДНК нуклеотиди в цьому сайті стають доступними для комплементарного спарювання з рибонуклеозидтрифосфатами. Взаємодію РНК-полімерази з ДНК- матрицею та ініціацію транскрипції зображено на рис 10.23.

Рис. 10.22. Структура бактеріального промотору
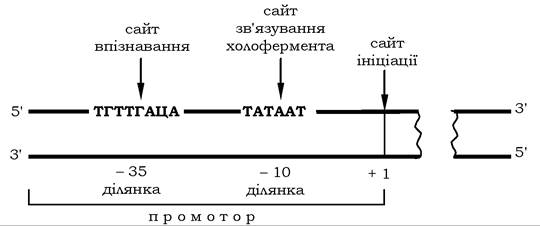
Рис. 10.23. Послідовність подій ініціації транскрипції РНК-полімеразою E. coli: N - будь-який нуклеотид
Для еукаріотичних організмів характерним є попереднє зв'язування з промотором низки білків, так званих загальних білкових факторів транскрипції. Назва "загальні" зумовлена присутністю цих факторів у всіх клітинах і абсолютною необхідністю їх для транскрипції більшості генів.
РНК-полімераза розміщує перший нуклеотид, причому обов'язково пуриновий (АТФ або ГТФ) із збереженням усіх трьох його фосфатних залишків. Потім утворюється перший 5',3'-фосфатний зв'язок з другим нуклеотидом.
У міру того як вивільняється промотор, до нього знову можуть приєднуватися нові молекули РНК-полімерази таким чином, що ген може транскрибуватися одночасно великою кількістю молекул ферменту.
Після того як синтезується олігонуклеотид із 8-10 нуклеотидних залишків, σ-субодиниця втрачає зв'язок з ферментом, а замість нього до РНК-полімерази приєднується декілька факторів елонгації.
Елонгація транскрипції. Фактори елонгації підвищують активність РНК-полімерази й полегшують розходження ланцюгів ДНК. Фермент послідовно каталізує реакцію приєднання 3'-ОН-групи нуклеотиду, розташованого в кінці зростаючого ланцюга до α-фосфату наступного рибонуклеозидтрифосфату. Утворення кожного фосфодіефірного зв'язку супроводжується вивільненням неорганічного пірофосфату, який швидко піддається гідролізу до неорганічного ортофосфату й таким чином робить реакцію енергетично вигідною.
Вибір чергового нуклеотиду в заново синтезуючому ланцюгу РНК визначається будовою комплементарного нуклеотиду в кодуючому ланцюгу ДНК таким чином, що азотисті основи А, Г, Т, Ц ланцюга ДНК кодують включення в ланцюг РНК відповідно основ У, Ц, А та Г.
Нарощування молекули РНК відбувається завдяки переміщенню РНК-полімерази вздовж ДНК шляхом приєднання чергового рибонуклеотиду, комплементарного тому дезоксирибонуклеотиду, який у даний момент перебуває в області активного центра РНК-полімерази.
У ході просування РНК-полімерази вздовж кодуючого ланцюга в напрямку 5'→3', попереду неї відбувається розходження, а позаду - відновлення подвійної спіралі ДНК. Приблизна швидкість руху ферменту й синтезу РНК - 30 нуклеотидів за секунду.
Термінація транскрипції відбувається після досягнення РНК-полімеразою на ДНК-матриці певних термінуючих ділянок (сайти термінації). На початку цих ділянок трапляються скупчення ГЦ-пар, локальна денатурація яких у ДНК проходить скрутно (порівняно з АТ-парами), що сприяє уповільненню просування РНК-полімерази й може служити сигналом до припинення транскрипції. До того ж ГЦ-збагачена ділянка може являти собою паліндром (інвертований повтор) - ділянка ДНК, обидва ланцюги якої мають однакову послідовність нуклеотидів при прочитуванні в різних напрямках. За наявності таких послідовностей розкручування подвійної спіралі ДНК у термінуючих ділянках створює умови для взаємодії між нуклеотидами одного ланцюга з утворенням "шпильки". Це полегшує відокремлення РНК від ДНК. Припиненню синтезу РНК сприяє й спеціальний білок - Rho-фактор. Він рухається вздовж ДНК услід за РНК- полімеразою, має роз'єднувальну активність і створює умови для дисоціації комплексу "ДНК - фермент - РНК".
Після завершення транскрипції та звільнення первинного транскрипту відновлюється двоспіральна структура ДНК. РНК- полімераза може вступати в наступний цикл транскрипції після приєднання субодиниці σ.
Транскрипція гена відбувається конвеєрним способом - з одним геном одночасно зв'язані декілька зростаючих РНК-транс- криптів. Це пов'язано з тим, що одна молекула РНК-полімерази проходить через промотор і просувається вздовж ДНК на деяку відстань, з промотором зв'язується наступна молекула ферменту й також починає транскрипцію. Тому на кожній транскрибуючій ділянці зазвичай присутні відразу декілька молекул ферментів, що рухаються одна за одною на відстані в 300-500 нуклеотид- них пар. Узагалі, середня відстань між ними залежить від "сили" промотору, зумовленої переважно транскрипційними факторами, і концентрацією РНК-полімерази.