БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 11. СТРУКТУРА І ВЛАСТИВОСТІ ВУГЛЕВОДІВ
11.3.Полісахариди
Полісахариди (поліози) є природними полімерами, побудованими з великої кількості залишків молекул моносахаридів та їхніх похідних. За моносахаридним складом полісахариди поділяють на гомополісахариди, або гомоглікани, які складаються із залишків моноз одного виду, і гетерополісахариди, або гетероглікани, молекули яких вміщують залишки різних моносахаридів. Полісахариди різняться також молекулярною масою та структурою молекул. За характером поліглікозидного ланцюга вони можуть бути лінійними та розгалуженими.
Через відсутність систематичної номенклатури полісахаридів застосовується раціональний принцип, згідно з яким назви полісахаридів походять від назви монози - мономера із заміною суфікса -оз на -ан. Наприклад, полісахариди, що складаються тільки із залишків D-глюкози, або D-фруктози називають відповідно D-глюканами або D-фруктанами. Якщо до складу поліози входять обидва види цих моноз, його називають D-глюко-D-фруктаном. Характер глікозидного зв'язку позначається буквами а- або β- попереду групової назви полісахариду: a-D-глюкани або β-D-фруктани.
Поряд із раціональними назвами найважливіших гомогліканів часто використовуються їхні давні емпіричні назви - крохмаль, клітковина, глікоген; або гетерогліканів - гепарин, пектин, гіалуронова кислота тощо.
Найпоширенішим мономером природних полісахаридів є D-глюкоза, серед інших моноз найчастіше зустрічаються D-маноза, D- і L- галактоза, L-арабіноза, D-ксилоза, D-глюкуронова кислота, D-галактуронова кислота, D-мануронова кислота, D-глюкозамін, D-галактозамін, сіалові кислоти.
Глікозидні зв'язки, що сполучають залишки моноз у молекулах полісахаридів, утворюються переважно за глікозидо-глікозидним типом, через що в молекулах поліоз практично немає вільних напівацетальних гідроксилів.
Полісахариди становлять основну масу тканин рослин, містяться в клітинах тварин, у мікроорганізмах, де виконують різноманітні важливі функції. Це, в першу чергу, механічні, насамперед опорні функції целюлози та її похідних у вищих рослин, водоростей та також хітину у ракоподібних, молюсків і комах. Полісахариди крохмаль, пектини, фруктозани у рослин і глікоген у тварин виконують резервну функцію, тобто акумулюють метаболічну енергію, яка може бути мобілізованою для процесів життєдіяльності. Захисні функції полісахаридів у тканинах тварин пов'язані з їхньою участю у створенні імунітету, захисного слизового шару епітелію шлунково-кишкового тракту та бронхолегеневих повітряних шляхів, синовіальної рідини суглобів тварин тощо. Серед багатьох різнобічних функцій поліоз слід виділити складні специфічні функції, які вони виконують у процесах згортання крові, нервової діяльності, взаємодії клітин у багатоклітинному організмі, взаємодії клітин і вірусів та ін.
За хімічним складом і будовою молекули полісахариди класифікують на групи:
✵ глюкани (крохмаль, глікоген, целюлоза, інші глюкани);
✵ манани (манани, глюкоманани, галактоманани);
✵ фруктани (фруктани, глюкофруктани);
✵ ксилани (ксилани, арабіноксилани, глюкуроноксилани);
✵ арабінани;
✵ галактани (галактани, арабіногалактани);
✵ поліуроніди (пектинові речовини, альгінова кислота);
✵ хітин;
✵ глюкозамінглікани.
Найважливішими природними глюканами є целюлоза, крохмаль, глікоген.
Целюлоза, або клітковина, є найпоширенішою у природі органічною сполукою, є основою клітинної оболонки рослинних організмів, бактерій, а також деяких нижчих тварин (Tunicata). Її присутність надає клітинам і тканинам цих організмів механічної міцності, пружності та еластичності. Це високомолекуляр- ний лінійний вуглевод, побудований із глюкозильних залишків сполучених між собою β-1,4-глікозидним зв'язком, а також водневими зв'язками між гідроксилами третього атома вуглецю монози та гетероатомом кисню сусіднього глюкозильного залишку:
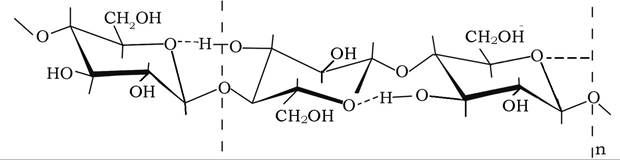
Целюлоза - біла аморфна речовина з характерною впорядкованою структурою. Макромолекулярні лінійні ланцюги її довжиною від 1500 нм за допомогою міжмолекулярних водневих зв'язків утворюють мікрофібрили, кожна з яких може містити по декілька десятків лінійних макромолекул. Мікрофібріли, у свою чергу, асоційовані у волокно, в якому осі мікрофібріл розміщені під кутом до осі волокна. Лінійні молекули целюлози на визначених ділянках волокон зібрані в упорядковані пучки, паралельні осі волокна, утворюючи кристалічні міцели довжиною ~60 нм і діаметром 5 нм, які мають велику механічну міцність та хімічну стійкість. Проміжні міжміцелярні аморфні ділянки волокна характеризуються значно меншою міцністю і в них швидко проходять реакції гідролізу, окиснення, заміщення тощо.
Складна структура рослинної клітинної стінки з різнобічною орієнтацією в ній мікрофібрил надає деревині, що складається з клітинних оболонок, тих механічних властивостей, завдяки яким вона посідає винятково важливе місце серед будівельних матеріалів, здавна використовуваних людиною.
Целюлоза нерозчинна у воді, ефірі, ацетоні, спирті й вуглеводнях. Вона стійка в розведених розчинах кислот і лугів, але розчиняється в концентрованих розчинах мінеральних кислот і органічних основ. У присутності концентрованих кислот целюлоза під
дається гідролізу з утворенням β-Ό-глюкози, що є основою виробництва гідролізного етилового спирту. Ферментативний гідроліз целюлози відбувається за участю ферментів бактерій-симбіонтів, присутніх в організмах жуйних тварин та комах (термітів).
Бактеріальні ензими целюлаза й целобіаза послідовно розчиняють молекулу целюлози на молекули целобіози та D-глюкози за схемою:
![]()
Хімічні властивості целюлози зумовлені наявністю в складі її молекули вільних гідроксильних груп, що сприяють утворенню алголятів, ефірів і естерів целюлози.
У промисловості у великий кількості виробляють алкаліцелюлози, метил-, етил-, бензилцелюлози, нітрати целюлози, а також ксантогенат- і ацетатцелюлози, які використовують у харчовій, косметичній, текстильній промисловості, та у виробництві паперу, лаків, плівок, пластмас, вибухових речовин тощо.
У біохімічних дослідженнях для розділення амінокислот, пептидів, білків, гормонів, нуклеотидів і нуклеїнових кислот методами іонообмінної хроматографії широко використовують діетиламіноетил- (ДЕАЕ) і карбоксиметилцелюлозу (КМЦ) як іонобмінники.
Крохмаль - основний резервний вуглевод рослин і важливий компонент харчування людини, є продуктом полімеризації глюкози, що утворюється шляхом фотосинтезу в зелених листках. Синтезується крохмаль у зернах злакових і бульбах овочевих, де й накопичується у значних кількостях (наприклад, у зернах рису - до 80 %, пшениці - до 85 %, у бульбах картоплі - 25 %) у вигляді мікроскопічних овальних зерен діаметром від 2 до 150 мкм. Крім глюкози до складу природного крохмалю входить білок (до 0,8 %), вищі жирні кислоти (0,6 %), а також фосфатна кислота (до 0,7 %).
Крохмаль - білий, без смаку й запаху порошок, нерозчинний у воді за кімнатної температури. При нагріванні у воді при 65-70 °С спостерігається клейстеризація крохмалю - його зерна набухають і розпадаються, утворюючи колоїдний розчин. Розчини крохмалю оптично активні, кут питомого обертання, наприклад картопляного крохмалю, [α]D20 = 204,3o.
Тривале нагрівання крохмалю у воді у присутності 10 % розчину H2SO4 спочатку приводить до декстринізації - утворення низькомолекулярних декстринів, згодом до ступінчастого гідролізу з утворенням олігосахаридів і D-глюкози за схемою:
Розчини крохмалю й декстринів при взаємодії з розчином йоду набувають синього, фіолетового, оранжевого та червоного забарвлення, що й використовується для ідентифікації цих сполук.
За хімічною будовою крохмаль є структурно гетерогенним гомополісахаридом, який складається з двох компонентів: амілози та амілопектину.
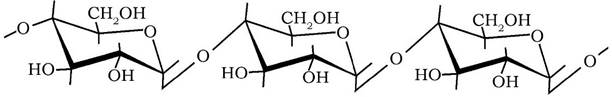
Амілоза має лінійний ланцюг, в якому глюкозильні залишки сполучені між собою а-1,4-глікозидним зв'язком:
![]()
Для амілози характерна відносно невелика молекулярна маса (20-60 кДа), висока в'язкість і нестабільність водних розчинів, утворення темно-синього забарвлення з розчином йоду. Вона легко гідролізується амілолітичними ферментами - а- і β-амілазами до молекул дисахариду мальтози:
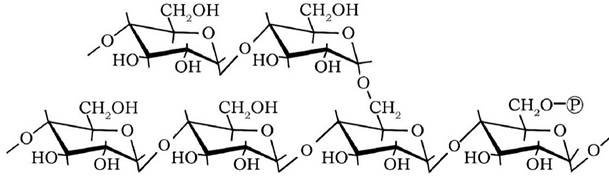
Амілопектин - розгалужений глюкан, у молекулах якого через кожні 20-25 ланок а-1,4-глікозідних зв'язків спостерігаються розгалуження, утворені а-1,6-глікозидним зв'язком. У середньому в амілопектині 16 % глюкозильних залишків фосфорильовані:

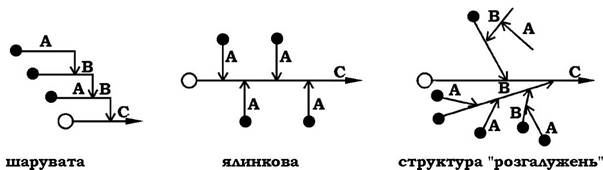
Порівняно з амілозою амілопектин має складнішу будову, яка складається з поліглюкозильних ланцюгів трьох типів: основного (С), бокового (А) і проміжного (В). Ці ланцюги можуть утворювати структури трьох видів: шарувату, ялинкову та структуру "розгалужень":
Амілопектин характеризується великою молекулярною масою (вище 103 кДа), важкою розчинністю у воді, нездатністю кристалізуватися. У воді він утворює високостабільні колоїдні розчини, які забарвлюються йодом у червоно-фіолетовий колір. Ферментативний гідроліз амілопектину відбувається повільніше порівняно з амілозою: β-амілаза послідовно відщеплює від нередукуючих кінців бічних поліглікозидних ланцюгів молекули дисахариду мальтози впритул до точки розгалуження і припиняє гідроліз перед останнім а-1,4-глікозидним зв'язком. Надалі продукти її гідролізу - високомолекулярні декстрини розщеплюються а-амілазою (до низькомолекулярних декстринів) і глюкоамілазою, яка гідролізує і а-1,3- і а-1,6-глікозидні зв'язки, відщеплюючи молекули D-глюкози від нередукуючих кінців молекул декстринів:

Мальтоза, у свою чергу, розщеплюється мальтозою (α -глюкозидазою) до двох молекул D-глюкози. Отже, за участю амілоліти- чних ферментів шлунково-кишкового тракту тварин крохмаль практично повністю розщеплюються до молекул D-глюкози.
У харчовій і текстильній промисловості широко використовують модифіковані крохмалі, тобто крохмалі, природні властивості яких змінені внаслідок фізичної, хімічної, біологічної та комбінованої обробки ультразвуком, іонізуючим випромінюванням, окисниками, кислотами, іншими хімічними реагентами, ферментами. У харчовій промисловості використовують окиснені, набухаючі, фосфатні, ацетильовані та інші модифіковані крохмалі для виробниц
тва хліба, пудингів, морозива й молочних концентратів, майонезів, соусів, м'ясних напівфабрикатів, продуктів дієтичного та дитячого харчування тощо. У текстильній галузі використовують заміщені крохмалі у виробництві натуральних і штучних волокон.
Глікоген є резервним вуглеводом у тканинах тварин і людини, де виконує роль тваринного крохмалю. Він синтезується практично в усіх клітинах організму, але особливо високий вміст його в печінці - до 7 % та у м'язах - до 0,9 % від сирої маси тканини. Глікоген міститься також у грибах, дріжджах, мікроорганізмах і деяких вищих рослинах (кукурудзі цукровій).
Чистий глікоген - білий аморфний порошок, який добре розчиняється у воді. Розчини глікогену опалесціюють, оптично активні, кут питомого обертання [α]D20 = 196°.
Глікоген є розгалуженим полісахаридом, що складається із залишків a-D-глюкопіраноз, які в лінійній частині молекули сполучені а-1,4-зв'язками, а в точках розгалуження а-1,6-глікозидним зв'язком, іноді а-1,3-зв'язком.
Порівняно з амілопектином молекула глікогену має більш високий ступінь розгалуження: а-1,6-зв'язок припадає на 8-12 ланок α-1,4-глікозидних зв'язків, завдяки чому глікоген компактніший за структурою, ніж амілопектин. Форма його макромолекули наближається до сферичної, молекулярна маса досягає 1 · 106 кДа (рис. 11.1).

Рис. 11.1. Схема будови макромолекули глікогену за У. Уїланом:
А - прямі бокові ланцюги, сполучені з іншими ланцюгами а-1,6-зв'язком; В - проміжні ланцюги, з яким сполучаються ланцюги А;
С - єдиний ланцюг у молекулі з вільним відновним кінцем
Макромолекулярна структура глікогену гетерогенна, складається з частинок трьох типів. Найбільші з них діаметром 60-200 мкм і молекулярною масою 105-106 кДа називають а-частинками; найменші, діаметром 20-40 мкм і молекулярною масою 2-5 ·103 кДа, - γ-частинками; і проміжні, що складаються з декількох γ-частинок, називаються β-частинками (рис. 11.2):
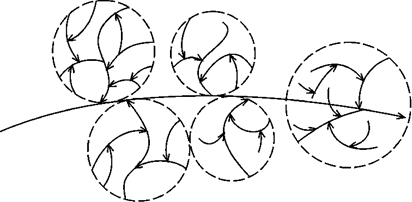
Рис. 11.2. Схема будови частинкового глікогену за Л. Лелуаром
Глікоген легко піддається кислотному або ферментативному гідролізу, під час якого розщеплюється з утворенням олігосахаридів - мальтотріози, ізомальтотріози, ізомальтози, мальтози та кінцевого продукту α-D-глюкози, що є ще одним свідченням сполучення глюкозильних залишків в молекулі глікогену за участю а-1,4- та а-1,6-глікозильних зв'язків.
Хітин - поширений у природі гомополісахарид, з якого побудовані кутикули комах або екзоскелети членистоногих (Arthropoda), а також покриття грибів (Eumycophyta), клітинних стінок багатьох мікроорганізмів (Aspergillus niger, Neurospora crassa та ін.) і деяких нижчих рослин. Хітин є лінійним полісахаридом з молекулярною масою 150-200 кДа, що складається із залишків N-ацетил- глюкозаміну, з'єднаних між собою В-1,4-глікозидними зв'язками:
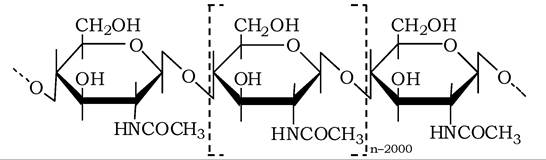
Хітин можна розглядати як 2-N-ацетилпохідне целюлози, він, як і целюлоза у рослин, у природі виконує механічну та захисну функцію в організмах, які його синтезують.
Гетероглікани Разом із целюлозою в утворенні клітинної стінки рослин беруть участь також інші полісахариди: геміцелюлози, пектинові речовини. Геміцелюлозами називають групу гетерополісахаридів, які розрізняються за моносахаридним складом і будовою основної й розгалуженої частини поліглікозидного ланцюга. Цю різноманітну за складом і структурою макромолекул групу утворюють манани, галактани, фруктани, ксилани, арабінани.
Манани містяться в деревині хвойних, водоростях і складаються із залишків D-манопіранози, які можуть бути сполучені β-1,3-, β-1,4-, β-1,6-глікозидними зв'язками.
Галактани побудовані залишками D-галактопіраноз за допомогою β-1,4-глікозидного зв'язку. Містяться у водоростях (агар), насінні й коренеплодах вищих рослин.
До фруктанів належить найпоширеніший у природі поліфруктозид інулін, що накопичується як резервний вуглевод у значній кількості в бульбах рослин родини складноцвітних (топінамбурі, артишоках, цикорію та ін.). У молекулі інуліну поряд з фруктофуранозильними залишками, сполученими β-2,1-глікозидним зв'язком, містяться залишки α-D-глюкопіранози (3-6 % від загальної кількості моноз). Молекулярна маса інуліну становить 5-6· 103 Да. Він добре розчиняється у воді, його розчини оптично активні ([α]D20 = -39°); кристалізується з розчинів у вигляді сферокристалів. Із трьох молекулярних форм інуліну (α-, β- і γ-) найбільш біологічно активний γ-інулін, який виявляє значну гіпоглікемічну, гіпохолестеричну та фібрінолітичну активність.
Пектинові речовини, які разом з геміцелюлозою є постійними супутниками целюлози, містяться у клітинних стінках і міжклітинній речовині всіх вищих рослин і водоростей. У деяких із них вони накопичуються в значний кількості - в яблуках до 15 %, у шкірці цитрусових - до 30 % від сухої маси речовини. У більшості рослин протягом певного часу пектинові речовини перебувають у водонерозчинній формі протопектину, який згодом при дозріванні плодів переходить у розчинну форму - пектин, який є складовою соків плодів і овочів.
Структурною основою молекул пектинових речовин є полімерний ланцюг із залишків D-галактуронової кислоти, з'єднаних між собою а-1,4-глікозидним зв'язком.
Поліози, які містять метилові ефіри D-галактуронової кислоти, називають пектиновими кислотами, а їхні солі - пектинатами:
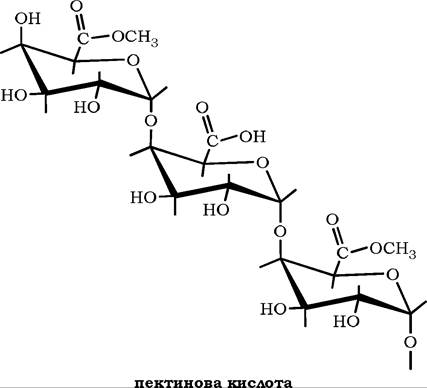
Неметаксильовані полігалактуроніди називають пектовими кислотами, а їхні солі - пектатами.
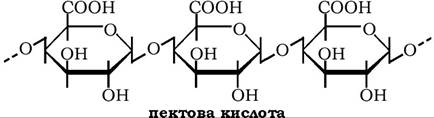
До складу молекул пектинів можуть також входити від 5 до 25 % нейтральних цукрів: L-арабіноза, L-рамноза, D-галактоза; у меншої кількості присутні D-ксилоза, L-фруктоза та їхні метилові ефіри.
Специфічною властивістю пектинових речовин є їхня здатність утворювати гелі, широко використовувані в харчовій промисловості у виробництві желе, джемів, мармеладів тощо, а також у медицині як ефективні сорбенти й комплексоутворювачі для детоксикації ендо- та екзотоксинів, виведення радіонуклідів і важких металів з організму людини.
Агар-агар - складний полісахарид, міститься в деяких водоростях північних морів, є сумішшю поліозагарози й агаропектину. Агароза побудована залишками D-галактопіранози та 3,6-ангідро-L-галактопіранози, почергово сполученими β-1,4- і а-1,3-глікозидними зв'язками:
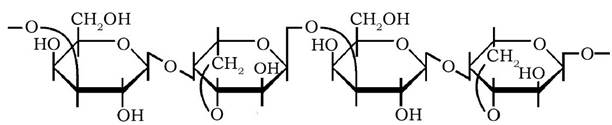
Агаропектин складається з D-галактозильних залишків, частина яких є сульфатованою.
При нагріванні у воді агар-агар утворює розчини, котрі при охолодженні перетворюються на гелі. Ці властивості агар-агару широко використовують у харчовій промисловості та біотехнології для приготування культуральних середовищ.
Вуглеводи сполучної тканини. Сполучна тканина, на яку припадає до 50 % маси всіх тканин організмів тварин, складається з міжклітинної (основної) речовини, клітинних елементів і волокнистої (колагенової) структури.
Основною речовиною є сильногідратований гель, що утворюється високомолекулярними сполуками - білками й вуглеводами, які становлять близько 30 % маси міжклітинної речовини. Вуглеводний компонент основної речовини представлений гетерополісахаридами - глюкозажінгліканажи (стара назва - мукополісахариди), важливою структурною ознакою яких є наявність дисахаридних одиниць, складених залишками гексуронових кислот і аміноцукрів.
Глюкозамінглікани є гідрофільними сполуками, які мають багато полярних ОН-груп і значний негативний заряд завдяки великій кількості карбоксильних і сульфогруп. Це зумовлює приєднання до них позитивно заряджених катіонів K+, Na+, Cu2+, Mg2+, що значно підвищує здатність глюкозамінгліканів утримувати воду.
У складі мономеру-дисахариду зазвичай присутні гексуронові кислоти: β-D-глюкуронова кислота та її епімер β-L-ідуронова кислота й аміноцукри. Найчастіше це D-глюкозамін і D-галактозамін та їхні ацетильні похідні — N-ацетил-D-глюкозамін і N-ацетил-D- галактозамін, сполучені між собою β-1,3-глікозидним зв'язком, за винятком мономерів гепарину, де має місце а-1,4-зв'язок.

За будовою мономерів, їхньою кількістю та характером зв'язку глюкозамінглікани поділяють на гіалуронову кислоту, хондроїтинсульфати, гепарансульфат і гепарин.
Гіалуронова кислота - біла, тверда, аморфна речовина, розчинна у воді й нерозчинна в органічних розчинниках. Оптично активна, кут питомого обертання у воді [α]D20 = -70° - 80o.
Гіалуронова кислота - єдиний несульфатований представник глюкозамінгліканів. Її молекули мають невпорядковану конформацію у вигляді хаотично згорнутих клубків молекулярної маси від 5·104 до 8·106Да. Побудовані вони з мономерів - дисахаридів, які складаються із залишків β-D-глюкуронової кислоти та N-ацетил-β-D-глюкозаміну, сполучених β-1,3-зв'язком. У свою чергу, мономери з'єднуються в молекулу лінійного полімеру за допомогою Р-1,4-глікозидного зв'язку:
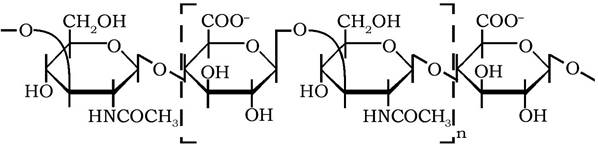
Завдяки іонізації карбоксильних груп залишків гексуронових кислот у її молекулах гіалуронова кислота має здатність утворювати комплекси з білками.
У тканинах і рідинах гіалуронова кислота існує у вільному стані або асоційована з білками, утворюючи дуже в'язки розчини. Найбільше її у склистому тілі, сухожиллях, шкірі, синовіальній рідині суглобів. У мезенхимних та інших тканинах вона забезпечує їхню стійкість до механічного стиснення, запобігає проникненню інфекції тощо.
Хондроїтинсульфати відрізняються між собою місцем розташування залишків сульфатної кислоти. У різних тканинах і органах тварин присутні чотири різновиди хондроїтинсульфатів, які позначаються літерами А, В, С і D.
Хондроїтинсульфати є у хрящах, шкірі, сухожиллях, клітинах серця, в аорті й артеріях, рогівці, кістках, склері, пупковому канатику та ін.
Дисахаридні фрагменти молекули хондроїтинсульфату А (хондроїтин-4-сульфату) містять залишки P-D-глюкуронової кислоти та К-ацетилгалактозамін-4-сульфату, сполучені Р-1,3-глікозидним зв'язком:
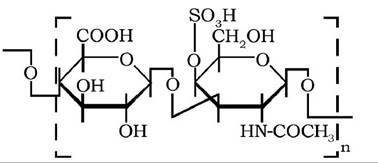
У молекулі хондроїтинсульфату С (або хондроїтин-6-сульфату) дисахаридні фрагменти складаються із залишків β-Ό-глюку- ронової кислоти й N-ацетилгалактозамін-6-сульфату також за допомогою β-1,3-зв'язку:

Головною відмінністю хондроїтинсульфату В (або дерматансульфату) від попередніх хондроїтинсульфатів є наявність у складі дисахаридної одиниці L-ідуронової кислоти замість D-глюкуронової:

Хондроїтинсульфат D, виділений із хрящової тканини безхребетних, відрізняється від інших хондроїтинсульфатів високим вмістом залишків сульфатної кислоти. У дисахаридних фрагментах цього хондроїтинсульфату сульфатовані залишки D-глюкуронової кислоти (біля С2 або С3) і N-ацетилгалактозаміну при С6.
Гепарансульфат і гепарин є також високосульфатованими глюкозамінгліканами молекулярної маси 15-20 кДа. До складу їхніх мономерів входять глюкуронат-2-сульфат і N-ацетилглюко-замін-6-сульфат, сполучені між собою а-1,4-глікозидним зв'язком:

Ці глюкозамінглікани добре розчиняються у воді, їхні розчини оптично активні ([α]D20 = 45 - 70o), а також мають найнижчу в'язкість серед усіх глюкозамінгліканів.
Гепарансульфат присутній на поверхні тромбоцитів і ендотеліальних клітин. Гепарин уперше був знайдений у печінці, але крім печінки високий вміст гепарину спостерігається на поверхні та всередині клітин у легенях, м'язах, і в меншій кількості - серці, тимусі, селезінці, нирках і крові. Гепарин і гепарансульфат мають антикоагулятивні властивості, тому їх широко використовують у медичній практиці як антикоагулянти.
Зазвичай у нативному стані глюкозамінглікани зв'язані з білком нейтральним трисахаридом галактозил-галактозил-ксилозою через залишок серину поліпептидного ланцюга:
![]()
Полісахаридні ланцюги сульфатованих глюкозамінгліканів мають спіральну конфігурацію. Довгі полісахаридні ланцюги глюкозамінгліканів утворюють глобули, які займають досить великий об'єм.
Молекули протеогліканів - складних білків міжклітинного матриксу складаються з особливого білка, названого СОК-білком, до котрого за участю трисахаридів приєднані молекули глюкозамінгліканів. До одного поліпептидного ланцюга СОК-білка може приєднатися до 100 поліглікозидних ланцюгів глюкозамінгліканів (рис. 11.3).
Спіральні ланцюги глюкозамінгліканів у складі молекул протеогліканів утворюють пружні макромолекулярні сітчасті структури з визначеними розмірами пор, які виконують функції молекулярного сита при перенесенні поживних речовин і продуктів метаболізму. Розміри пор детермінуються формою глюкозамінглікану, який домінує в даній тканині. Разом із гіалуроновою кислотою й особливими зв'язувальними білками протеоглікани беруть участь у створенні складних надмолекулярних структур (рис. 11.4):

Рис. 11.3. Схема будови макромолекул протеогліканів
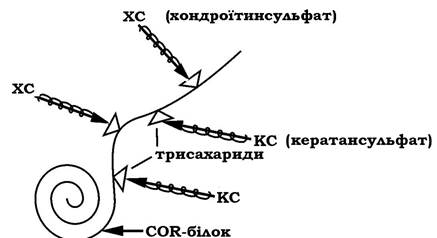
Рис. 11.4. Будова надмолекулярної структури основної речовини міжклітинного матриксу
Крім протеогліканів білковий компонент основної речовини міжклітинного матриксу включає глікопротеїни, вуглеводна частина яких дуже варіабельна. Глікопротеїни - це складні білки, вуглеводна частина яких представлена лінійними або розгалуженими олігосахаридами, що складаються, як правило, із залишків глюкози, галактози, манози, метилманози, ксилози, арабінози, рамнози, фікози, глюкозаміну й галактозаміну. На кінцях олігосахаридних ланцюгів молекул глікопротеїнів зазвичай розташовані залишки сіалових кислот. Вуглеводний компонент у молекулах глікопротеїнів може становити від 1 до 60 % маси цих білків. До глікопротеїнів відносять більшу частину білків міжклітинного матриксу і плазми крові, а також білки, розташовані на зовнішній поверхні клітинних мембран тварин.
До глікопротеїнів міжклітинного матриксу належить, зокрема, фибронектин (молекулярної маси 440 кДа). Його молекули складаються з двох поліпептидних ланцюгів, з'єднаних дисульфідним містком, і вуглеводного компонента, який має центри зв'язування з протеогліканами, з колагеновими та еластиновими волокнистими структурами матриксу, а також з гліколіпідами клітинних мембран. Таким чином, фибронектин виконує функцію "молекулярного клею», тобто сприяє адгезії та зв'язуванню клітин між собою.
До мембранних глікопротеїнів відноситься також глікофорин, присутній у клітинній мембрані еритроцитів (рис. 11.5). Глікофорин ("переносник цукру") є інтегральним мембранним білком (молекулярної маси 30 кДа), що містить 130 амінокислотних залишків і вуглеводний компонент, на який припадає приблизно 60 % маси всієї молекули. Глікофорин вбудований у мембрану таким чином, що гідрофільний С-кінець його поліпептидного ланцюга, складений залишками глутамінової та аспарагінової кислот і при рН 7,0 є негативно зарядженим, знаходиться в цитоплазмі, середня гідрофобна частина білка проходить крізь мембрану, а N-кінець поліпептиду виходить на зовнішню поверхню клітинної мембрани. До нього N- і О-глікозидними зв'язками приєднаний полярний вуглеводний компонент, котрий включає до 16 олігосахаридних ланцюгів, які містять групові антигенні детермінанти, що визначають ту чи іншу групу крові (А, В, О), а також беруть участь у рецепції вірусів грипу.
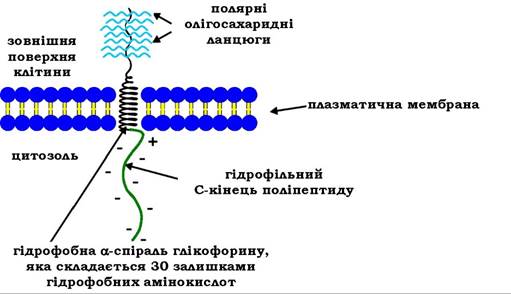
Рис. 11.5. Схема розташування глікофорину в плазматичній мембрані еритроцита
Ряд глікопротеїнів плазми крові формують антипротеїназну систему крові, зокрема, це а-1-антитрипсин, інтер-α-інгібітор трипсину, термокислотостабільний інгібітор трипсину, α-1-ан- тихімотрипсин тощо; частка вуглеводів у них становить 20-35 % молекулярної маси.
Усі імуноглобуліни, або антитіла, сироватки крові, а саме: IgA, IgG, IgM, IgD, IgE є глікопротеїнами, що містять ковалентно зв'язані олігосахаридні ланцюги, які становлять від 3 до 13 % маси цих макромолекул.
Таким чином, олігосахаридні ланцюги в складі молекул глікопротеїнів беруть безпосередню участь в утворенні основної речовини міжклітинного матриксу, у процесах міжклітинних взаємодій, рецепції антигенів, вірусів, імунологічної та антипротеїназної властивостях крові тощо.