БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 14. МЕТАБОЛІЗМ ЛІПІДІВ
14.6. Метаболізм фосфоацилгліцеролів
Для фосфоліпідів, як і для інших структурних компонентів біомембран, характерним є постійне оновлення, тобто стан динамічної рівноваги між їхнім синтезом і катаболізмом. Фосфоліпіди інтенсивно оновлюються в активно функціонуючих тканинах: печінці, нервовій тканині, ендо- та екзокринних залозах тощо.
Попередниками фосфоацилгліцеролів, як і триацилгліцеролів, є гліцерол-3-фосфат і КоА-ефіри жирних кислот, і в процесах утворення цих ліпідів існує декілька спільних етапів, котрі каталізуються одними ферментами (рис. 14.9, реакції 1-2):

Рис. 14.9. Схема біосинтезу фосфоацилгліцеролів
У клітинах тварин існує декілька шляхів утворення фосфоацилгліцеролів. Головними з них є синтез de novo і шлях реутилізації холіну.
Синтез de novo, відомий як цитидиновий шлях, здійснюється за участю цитидинових коферментів (рис. 14.9, реакції 3-6). Спочатку в реакції між фосфатидатом і цитидинтрифосфатом (ЦТФ) утворюється ЦДФ-діацилгліцерол, який далі реагує з гідроксилом аміноспирту (холіну, етаноламіну) або амінокислоти (серину, треоніну) з утворенням фосфоадилгліцеролів:
Слід зазначити, що цитидилові нуклеотиді виконують у цьому синтезі функцію, аналогічну функції уридилових нуклеотидів у синтезі глікогену. В обох випадках утворюються активовані проміжні метаболіти - УДФ-глюкоза та ЦДФ-діацилгліцерол.
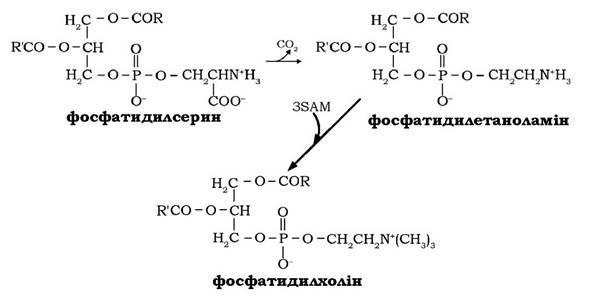
Декарбоксилюванням серинового залишку в молекулі фосфатидилсерину утворюється фосфатидилетаноламін. Фосфатидил холін синтезується метилюванням залишку етаноламіну, який входить до складу фосфатидилетаноламіну, за участю трьох молекул S-аденозилметіотну (SAM), який є універсальним донором метильних груп:

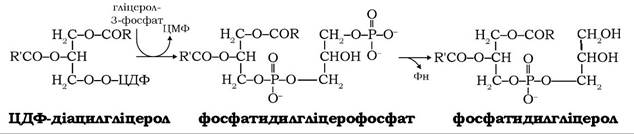
Інші мембранні фосфоліпіди - фосфатидилгліцериди, кардіоліпіни та фосфоінозитиди також синтезуються через ТТДФ- діацилгліцерол (рис. 14.9, реакції 7-11). Спочатку в реакції ЦДФ-діацилгліцеролу з гліцерол-3-фосфатом утворюється фосфатидил- гліцерофосфат, який далі дефосфорилюється й перетворюється у фосфатидилгліцерол:
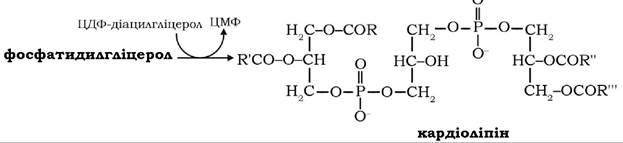
При взаємодії фосфатидилгліцеролу з ЦДФ-діацилгліцеролом синтезуються кардіоліпіни - головні фосфоліпіди мітохондріальних мембран:
При біосинтезі фосфоінозитидів фосфатидиловий залишок із молекули ЦДФ-діацилгліцеролу переноситься на інозитол з утворенням монофосфоінозитолу:
Монофосфоінозитиди перетворюються в ди- та поліфосфоінозитиди фосфорилюванням вільних гідроксилів інозитолу за участю АТФ.
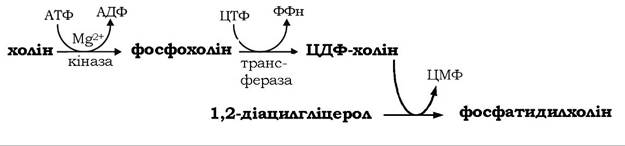
Ще один шлях синтезу фосфоацилгліцеролів (рис. 14.9, реакції 12-15) використовує як субстрат-попередник 1,2-діацилгліцерол, до якого приєднується активований фосфоетаноламін. Останній до цього активується етаноламінкіназою за участю двох нуклеотидтрифосфатів - АТФ і ЦТФ:

Фосфохоліновий залишок ЦДФ-холіну в наступній реакції переноситься на 1,2-діацилгліцерол, у результаті утворюється фосфатидилхолін.
У тварин, обмежених у можливості синтезувати лецитин de novo унаслідок нестачі метіоніну в їхньому звичайному раціоні, шлях реутилізації холіну є справді рятівним. Холін, який надходить в організм цих тварин з їжею, можна вважати додатковим вітаміном.
Розпад фосфоацилгліцеролів здійснюється гідролітичним шляхом за участю фосфоліпаз-ацилгідролаз (див. рис. 14.2).
Панкреатичні фосфоліпази й ферменти інших тканин характеризуються позиційною специфічністю і стереоспецифічністю до зв'язків, що гідролізують. Вони виявляють високу субстратну специфічність до різних груп фосфоліпідів (фосфоліпази С). Активність деяких фосфоліпаз (фосфоліпази А2 і D) регулюється іонами Са2+.
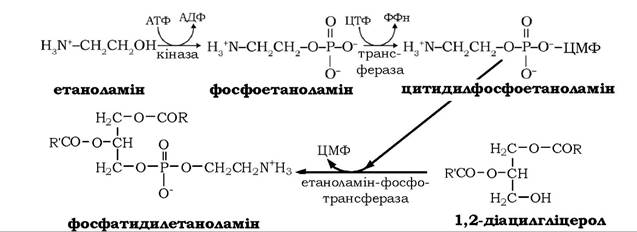
Аналогічним шляхом у тканинах тварин синтезується також фосфатидилхолін. Альтернативний шлях біосинтезу фосфатидил- холіну називають шляхом реутилізації, або "рятівним" (salvage) шляхом (рис. 14.9, реакції 16-18). На цьому шляху холін, який вивільняється при розщепленні фосфатидилхоліну та інших хо- ліновмісних сполук, фосфорилюється холінкіназою за участю АТФ з утворенням фосфохоліну, котрий далі реагує з ЦТФ, утворюючи активоване похідне холіну - ЦДФ-холін: